
2023-01-18 来源 : 学术查
大家都知道高速的糖酵解是肿瘤细胞的典型特征,肿瘤细胞通过糖酵解途径快速产生大量能量满足其高生物合成和生物能量的需求,以及维持其氧化还原状态,促进自身的生长发育,这也被称为Warburg效应。但是今天给大家介绍的这篇来自Science的文章,讲述的不是肿瘤细胞的糖酵解,而是发现了肿瘤细胞能够通过抑制免疫细胞糖酵解以逃避免疫杀伤促进自身的生长发育。这项研究为糖酵解与肿瘤细胞免疫逃逸提供了一个新的研究思路。具体内容如何,一起来看看吧。
研究背景
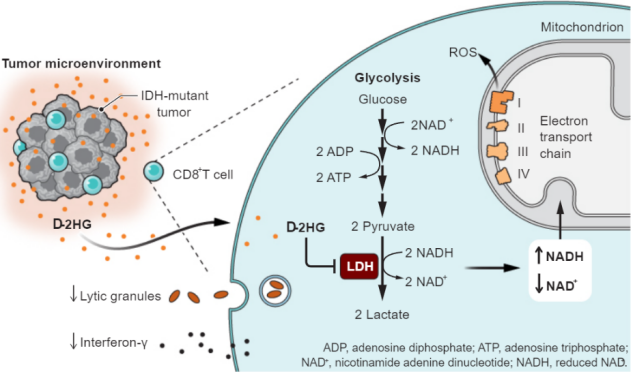
2022年9月30日,哈佛大学的Marcia C.Haigis团队在Science(IF:63.832)上发表题为“Oncometabolite D-2HG alters T cell metabolism to impair CD8+ T cell function”的文章。该研究发现肿瘤代谢物D-2HG可以抑制CD8+T细胞糖酵解,使得细胞增殖、细胞因子产生以及细胞毒性均被抑制,导致CD8+T细胞的抗肿瘤作用受到损害。该研究还提出了D-2HG促肿瘤的新机制,通过抑制乳酸脱氢酶(LDH)的活性,导致糖酵解通量和NAD(H)平衡的改变,使ROS水平升高,进一步抑制CD8+T的杀伤能力。


编码关键代谢酶的基因突变的发现表明代谢改变和疾病之间存在直接联系。而三羧酸循环中的富马酸水合酶、琥珀酸脱氢酶和异柠檬酸脱氢酶(IDH)突变的发现表明线粒体代谢物的积累会促进肿瘤的发生发展,这类代谢物往往被称为肿瘤代谢物(oncometabolites)。肿瘤代谢物除了在细胞内固有的功能外,还能够作用于肿瘤微环境,影响细胞间的相互交流。D-2-羟基戊二酸(D-2-hydroxyglutarate D-2HG)作为人异柠檬酸脱氢酶(IDH)功能获得性突变产生的一种肿瘤代谢物,通过改变肿瘤细胞的遗传途径,使得它们转变为更具侵袭性,快速分裂的状态,从而帮助肿瘤细胞的生长。然而很少有研究D-2HG如何影响肿瘤微环境中的其他细胞,包括CD8+T——通过释放颗粒酶和细胞因子达到杀伤肿瘤细胞的免疫细胞。
研究内容
1. D-2HG以急性且可逆的方式抑制CD8+ T细胞增殖、细胞毒性和IFN-γ信号
为了评估D-2HG影响抗肿瘤免疫的肿瘤细胞非自主机制,作者首先检测了肿瘤代谢物D-2HG和其异构体L-2HG对CD8+ T细胞功能的影响。通过不同浓度的共培养实验,确定了一个对细胞无毒害且易被吸收的作用浓度。研究发现,在相同的浓度下,D-2HG表现出更强的T细胞增殖抑制作用,并且随着D-2HG的移除能够缓解这种增殖阻滞,表明这种抑制作用是可逆的。CD8+T细胞通过分泌含有颗粒酶等细胞毒性蛋白分解靶细胞是T细胞杀伤肿瘤细胞的重要方式之一。作者研究发现D-2HG会导致CD8+T细胞的“脱粒”反应受损,并且INF-γ的产生和分泌也会受到显著抑制。体外杀伤实验也进一步证明了D-2HG会快速且可逆地破坏CD8+T细胞对肿瘤细胞的杀伤能力。
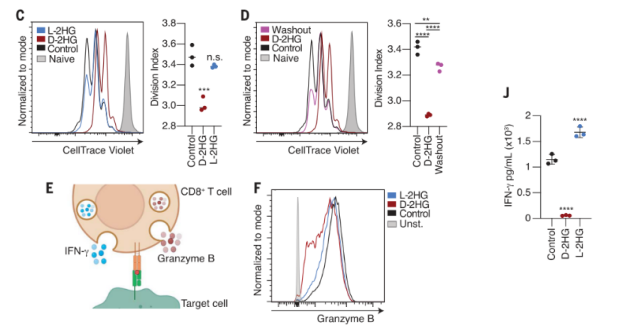
图1.(C)D-2HG抑制CD8+ T细胞增殖;(D)D-2HG的移除缓解了CD8+ T细胞增殖的抑制;(E-F)D-2HG抑制CD8+ T细胞颗粒酶B的产生和释放;(J)D-2HG抑制CD8+ T细胞IFNγ的产生和释放。
2. D-2HG通过损害LDH活性,降低NAD+/NADH比值,抑制CD8+T细胞的糖酵解
鉴于这种急性且可逆的特性与其作为表观遗传调节剂的既定作用无关,作者认为D-2HG可能通过影响T细胞代谢进而抑制其活性。作者首先测试了D-2HG和L-2HG是否会被T细胞代谢,通过WB分析和同位素示踪实验发现2HG不易被代谢。进一步的稳定代谢组学分析发现D-2HG的处理会促进糖酵解的中间产物的积累(如丙酮酸的增多)和终产物的减少(如乳酸的减少)。进一步的研究发现D-2HG能够显著性抑制CD8+T细胞糖酵解,而L-2HG则没有这一功能。并且研究发现这种对T细胞糖酵解功能的抑制是独立于HIF-1α途径的。
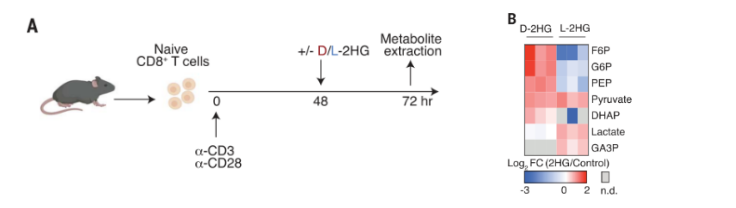
图2.(A)评估D/L-2HG对活化CD8+ T细胞稳态代谢产物水平急性(24小时)影响的实验设计示意图;(B) D/L-2HG对糖酵解影响的热图分析。
通过前面的研究发现D-2HG的处理导致T细胞中的丙酮酸含量升高,乳酸含量下调,这就提示D-2HG可能通过影响丙酮酸到乳酸的转化进而抑制T细胞糖酵解。在丙酮酸与乳酸的相互转变中,乳酸脱氢酶(LDH)发挥着至关重要的作用。作者接下来便检测乳酸脱氢酶活性是否会受到D-2HG的抑制。通过使用抗霉素A限制线粒体产生ATP以提高细胞对糖酵解的依赖性,作者发现在D-2HG存在的情况下LDH功能会受到损害。作为胞质NAD+再生的关键酶,LDH功能的损伤会进一步导致糖酵解通量和NAD+/NADH平衡的改变。
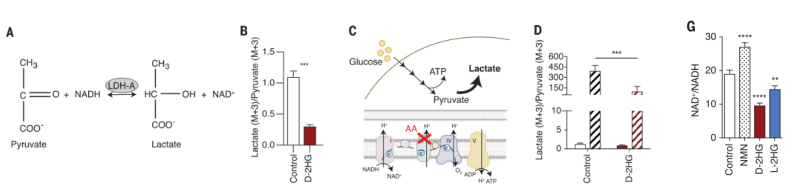
图3.(A) LDH反应示意图;(B)13C6葡萄糖与D-2HG或对照共处理24小时后CD8+ T细胞内乳酸(M+3)/丙酮酸(M+3)的比值;(C)抗霉素A (AA)处理对葡萄糖分解代谢的代谢影响示意图;(D)经过短时间的D-2HG预处理,然后抗霉素A(AA)和13C6葡萄糖共处理的细胞内乳酸(M+3)/丙酮酸(M+3)比值;(G)与对照,1 mM烟酰胺单核苷酸(NMN),20 mM D-2HG或20mM L-2HG共处理24h的CD8+ T细胞的NAD+/NADH比值。NMN提高细胞内NAD+水平,因此用作阳性对照。
2. D-2HG的处理诱导CD8+T细胞的线粒体呼吸最大化,并增加细胞对线粒体复合物I的依赖性,导致了ROS的大量升高
据Erika L Pearce报道T细胞一旦被激活,其能量产生的方式便会由氧化磷酸化途径转变为糖酵解途径。通过Seahorse XF 细胞能量代谢分析发现D-2HG会使T细胞更加依赖于氧化磷酸化途径产生ATP。进一步研究发现D-2HG的处理导致线粒体膜电位的极化,耗氧率增加和胞外酸化率降低,以一种急性且可逆的方式增加线粒体呼吸。D-2HG的处理导致NADH含量的升高进一步增加了细胞对线粒体复合物I的依赖,导致ROS产生增加。作者随后发现对LDH的单一抑制便足以重复D-2HG对CD8+ T细胞代谢和增殖、IFN-γ信号通路和细胞毒性的影响。
最后作者检测了IDH突变的人肿瘤中CD8+T细胞的变化,发现其特征与D-2HG处理的小鼠CD8+T细胞特征相似。

图4.(A)D-2HG处理24小时后,线粒体氧化磷酸化和糖酵解对ATP产生的影响;(B)20 mM D-2HG处理24小时后对基础呼吸的影响;(O)鱼藤酮(Rotenone)靶点示意图;(P)1nM鱼藤酮和20mM D-2HG处理对细胞分裂的影响;(Q)通过CellROX染色对经20 mM D-2HG处理或不处理1天的CD8+ T细胞内ROS水平进行量化评估。以过氧化氢(H2O2,100 mM)和10 mM NAC分别作为阳性和阴性对照。
研究总结
这项研究强调了代谢物对肿瘤微环境的作用,为今后肿瘤免疫逃逸相关研究提供了一个新的思路——肿瘤代谢物对微环境中免疫细胞的作用。正如作者Marcia Haigis说:“我们的研究强调了这种癌症中的免疫成分,这在以前没有得到充分认识。我们现在知道,肿瘤细胞产生的代谢物可以影响附近的免疫细胞,使周围环境对癌症的敌意降低。”
图片来源:Science.2022;377(6614):1488-1489.
此外,对D-2HG产生的这种急性且可逆的免疫细胞代谢重编程与肿瘤细胞表观遗传变化之间的联系仍需更进一步探究,以及D-2HG对肿瘤细胞糖酵解潜在影响也需进一步论述。Marcia Haigis团队的这项工作提示着肿瘤细胞以非自主方式实现免疫逃逸的机制可能远不止此,不同代谢物对不同免疫细胞如NK细胞,T细胞等可能产生不同的影响,相信随着研究技术的不断更新,人们会对肿瘤逃逸机制的理解不断加深。而且这项工作也为今后肿瘤治疗提供了新的思路——通过研发阻断异柠檬酸脱氢酶IDH突变药物抑制肿瘤免疫逃逸。
版权声明:本网站所有注明来源“医微客”的文字、图片和音视频资料,版权均属于医微客所有,非经授权,任何媒体、网站或个人不得转载,授权转载时须注明来源:”医微客”。本网所有转载文章系出于传递更多信息之目的,且明确注明来源和作者,转载仅作观点分享,版权归原作者所有。不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。 本站拥有对此声明的最终解释权。

































 关注公众号
关注公众号 安卓客户端
安卓客户端










发表评论
注册或登后即可发表评论
登录注册
全部评论(0)