
2022-11-09 来源 : 学术查
一、引言
自噬是一种溶酶体依赖性的用于降解细胞器和蛋白质的分解代谢过程,其中形成的自噬体需与溶酶体结合形成自噬溶酶体才能发挥作用。
外泌体是细胞内另一种消除异常蛋白质的途径之一,当细胞处于应激状态时,外泌体能作为桥梁提醒邻近细胞并引起邻近细胞的响应。
而自噬和外泌体两者都是基金中标数较多的科研热点,对两者的机制及两者之间的关系进行研究无疑是非常具有价值和意义的。
二、研究背景
氧化锌纳米颗粒(ZnONPs)由于具有美白性能和较强的紫外光吸收能力,而被广泛应用于各种化妆品中如护肤霜和防晒霜。然而,氧化锌纳米颗粒的生物安全问题和潜在危害目前仍具争议,其被发现在中波紫外线(UVB)照射或过敏性皮肤模型中可能导致皮肤毒性。紫檀芪(PT)因其抗炎和自噬诱导能力被应用于多种疾病的治疗中,据报道其能有效地预防皮肤损伤。鉴于此,作者想要探究UVB和ZnONPs共同造成皮肤损伤的潜在机制以及PT的保护作用。
三、研究成果
中国台湾国立成功大学医学院的王應然教授课题组从事环境与毒理学研究,2022年1月5日该课题组在Particle and Fibre Toxicology(IF=9.112) 杂志上发表题为"Skin damage induced by zinc oxide nanoparticles combined with UVB is mediated by activating cell pyroptosis via the NLRP3 inflammasome-autophagy-exosomal pathway"的文章。作者发现氧化锌纳米颗粒在紫外线照射下会导致皮肤损伤,这一过程涉及炎症小体、焦亡、自噬功能障碍及外泌体的相互作用。而PT则通过调节炎症-自噬-外泌体途径来缓解皮肤损伤。研究表明ZnONPs造成皮肤损伤的机制,为其在化妆品中的应用提供了指导意见,并为理解自噬与外泌体两种途径之间的关系提供新的角度。


四、研究过程与结果
1.ZnONPs的物理性质和细胞毒性
作者首先通过透射电子显微镜和动态光散射等方法观察并测定ZnONPs的物理性质(图1)。作者使用ZnONPs和UVB共同处理人永生化角质形成细胞,发现角质形成细胞(HaCaT)出现膨胀和内容物流出等现象,暗示其可能发生了细胞焦亡(图2A)。细胞毒性实验也表明ZnONPs和UVB共同处理后,角质形成细胞的细胞活性显著降低,并且是ZnONPs浓度依赖性的(图2B)。使用PT处理后,能够保护角质形成细胞免受ZnONPs和UVB共同处理造成的细胞损伤(图2C)。以上结果表明,ZnONPs和UVB共同处理会对角质形成细胞造成严重的细胞损伤,而PT能够保护细胞免受这种损伤。
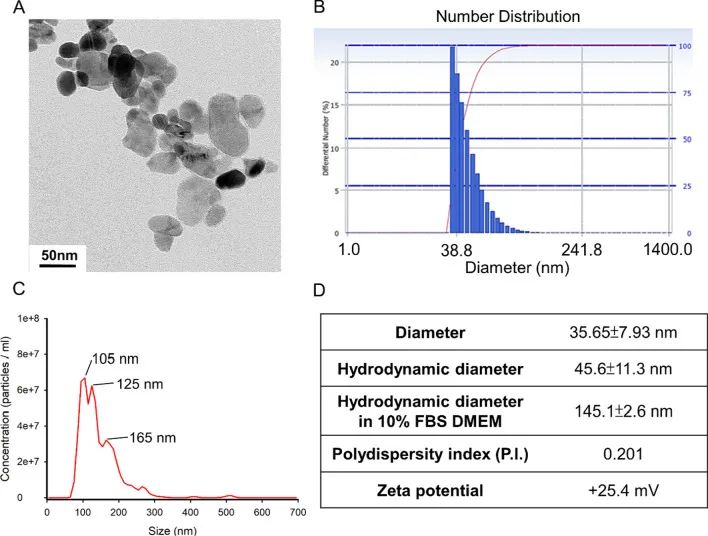
图1. ZnONPs的物理性质
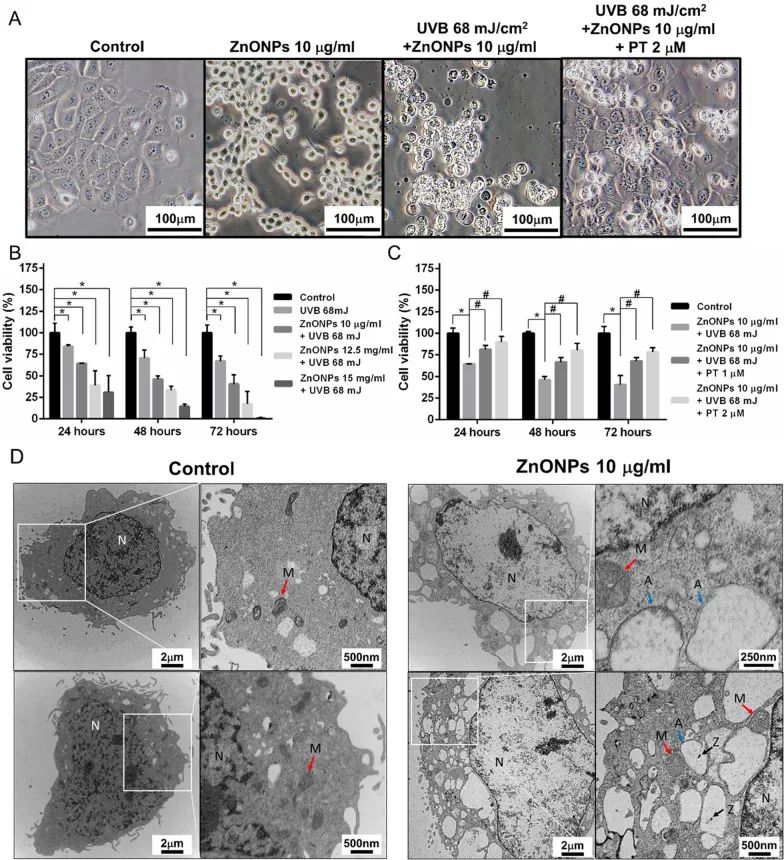
图2. ZnONPs和UVB共同处理后HaCaT细胞形态及活力变化
2.ZnONPs和UVB共处理诱导
线粒体损伤
据报道,ZnONPs引起细胞损伤可能是通过引起活性氧(ROS)水平升高和炎症反应造成的。而线粒体功能障碍通常是活性氧升高和炎症小体NLRP3激活的上游事件。作者使用透射电镜观察ZnONPs和UVB共同处理后HaCaT细胞内线粒体的超微结构,发现其线粒体外膜肿胀、嵴高度退化,表明线粒体损伤严重(图2D)。同时,ZnONPs和UVB共同处理后,HaCaT细胞线粒体膜电位显著降低,而细胞内ROS和线粒体ROS水平都显著上升,PT处理抑制了这些现象(图3A-F)。以上结果表明,ZnONPs和UVB共同处理会造成角质形成细胞线粒体损伤及ROS水平升高。
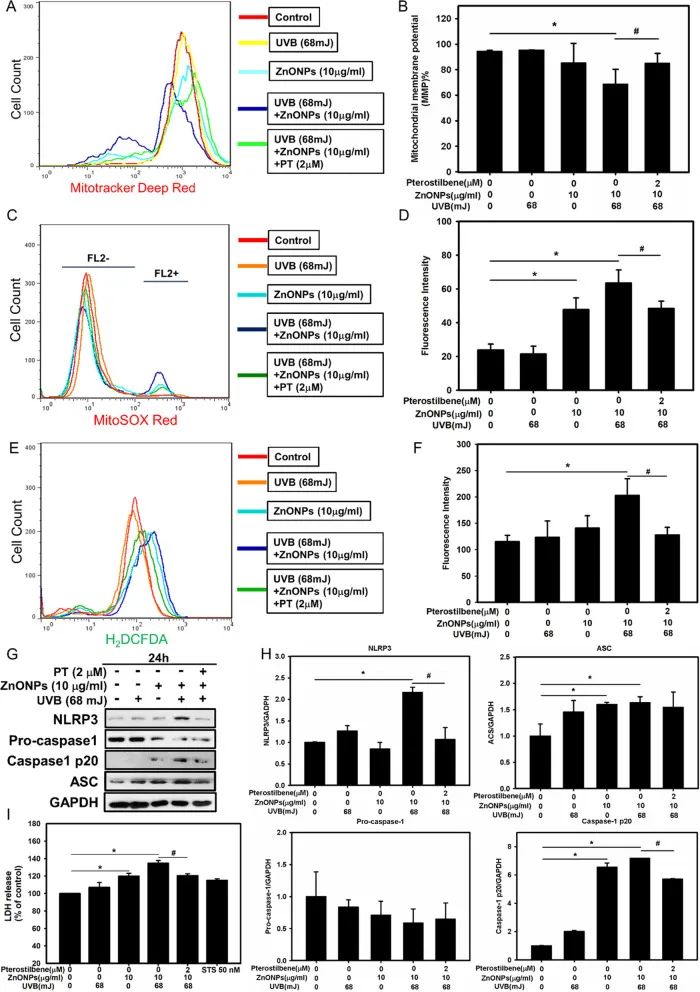
图3. ZnONPs和UVB共处理诱导线粒体损伤
3.ZnONPs造成的线粒体损伤能诱导
炎症和细胞焦亡
活性氧升高能激活NLRP3等炎症小体。作者检测了NLRP3相关蛋白NLRP3、ASC、caspase1等蛋白表达量的变化,发现ZnONPs和UVB共同处理会导致这些蛋白表达量上调(图3G-H)。基于caspase-1在HaCaT细胞中激活,作者假设可能发生了细胞焦亡。通过乳酸脱氢酶(LDH)释放量检测、流式细胞分析、以及Western blotting和免疫荧光检测焦亡相关因子(GSDMD-NT、caspase4、caspase5),发现ZnONPs和UVB共同处理后HaCaT细胞发生了细胞焦亡(图3I,图4)。而PT处理后则显著抑制了HaCat细胞的炎症和焦亡发生。以上结果表明,ZnONPs和UVB共同处理诱导的线粒体损伤能激活NLRP3炎症因子和细胞焦亡,而PT处理能够抑制这一现象。
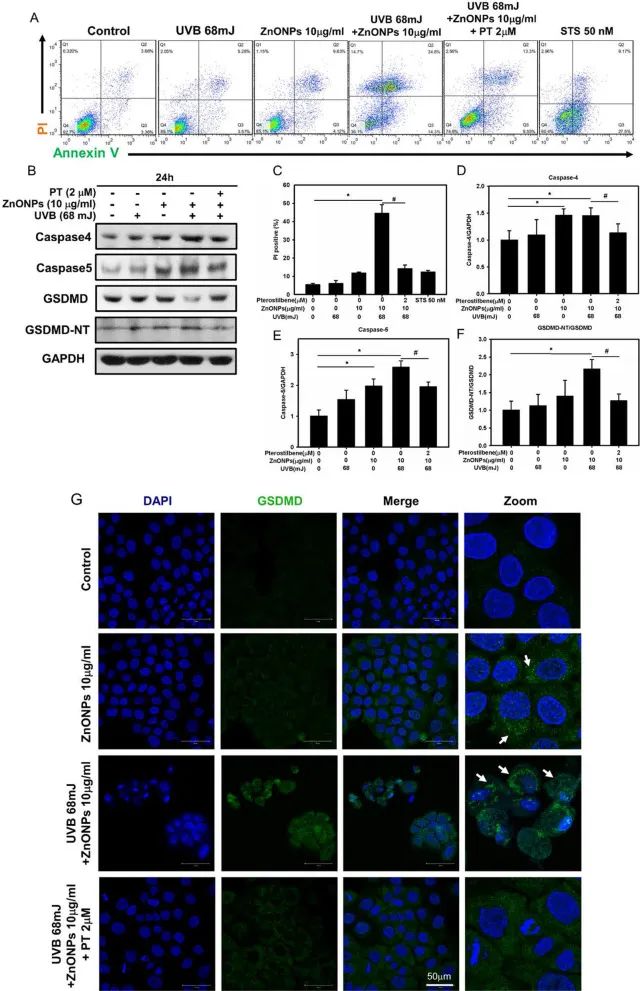
图4. ZnONPs和UVB共同处理触发炎症和焦亡
4.ROS在ZnONPs诱导的焦亡中
起关键作用
使用ROS抑制剂N-乙酰半胱氨酸(NAC)预处理HaCaT细胞,再使用ZnONPs和UVB共同处理HaCaT细胞,NAC能显著抑制ROS的产生(图5A-B)。而在ROS产生减少后,HaCaT细胞中PI与Annexin V双阳性的细胞比例、LDH的释放及炎症与焦亡相关因子的表达量都显著降低,表明细胞焦亡受到了抑制(图5C-H)。以上结果表明,ROS在ZnONPs和UVB共同处理导致的角质形成细胞NLRP3炎症小体激活与焦亡发生中起到关键作用。
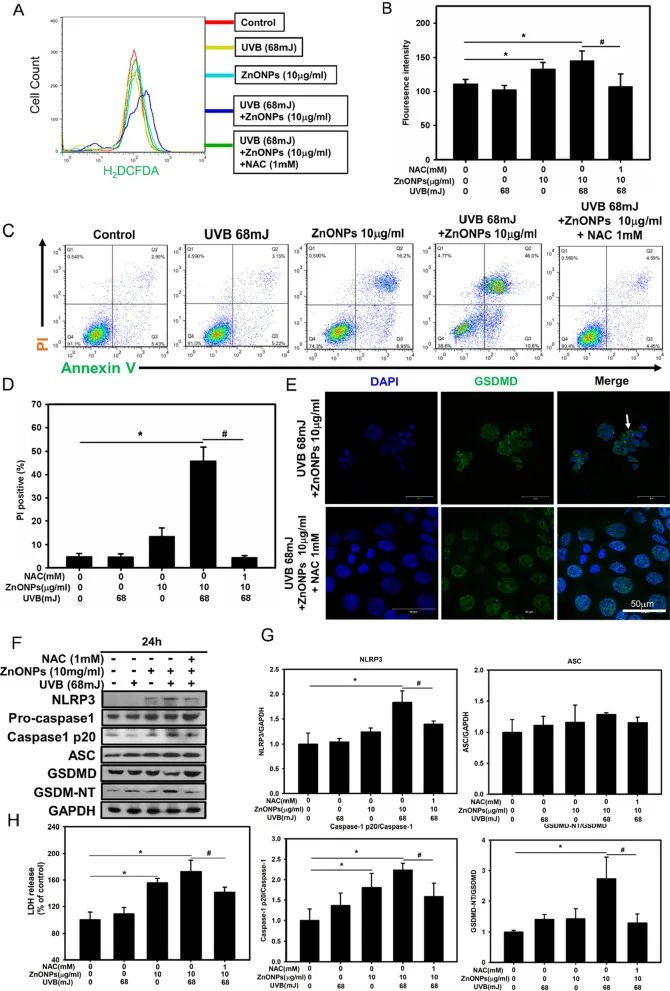
图5. ZnONPs诱导的NLRP3激活和焦亡由ROS介导
5.ZnONPs和UVB诱导自噬功能障碍
自噬激活可以通过自噬体与溶酶体结合形成自噬溶酶体降解细胞内的NLRP3炎症小体,而自噬功能障碍则会导致炎症小体的过度积累。在透射电镜下观察到ZnONPs和UVB共同处理的HaCaT细胞中出现大量自噬体,并且其中包含有ZnONPs(图2D)。通过吖啶橙(AO)染色及自噬通量蛋白LC3B和p62检测,发现ZnONPs和UVB共同处理的HaCaT细胞中AO荧光强度升高、LC3B及p62表达量显著上调,这提示细胞内的自噬通量受到阻断,自噬体无法与溶酶体结合形成自噬溶酶体,导致自噬功能障碍(图6A-D)。
为了进一步验证,作者使用LAMP-1(溶酶体标志物)和LC3B(自噬体标记物)免疫双荧光染色来观察溶酶体和自噬体的共定位,结果表明ZnONPs和UVB共同处理的HaCaT细胞中没有发生红色荧光和绿色荧光的共定位,说明溶酶体和自噬体无法融合;而PT处理组中则出现明显的黄色荧光,表明自噬溶酶体成功形成(图6E)。以上结果表明,ZnONPs和UVB共同处理会导致角质形成细胞的自噬功能障碍,而PT处理则能够恢复其自噬功能。
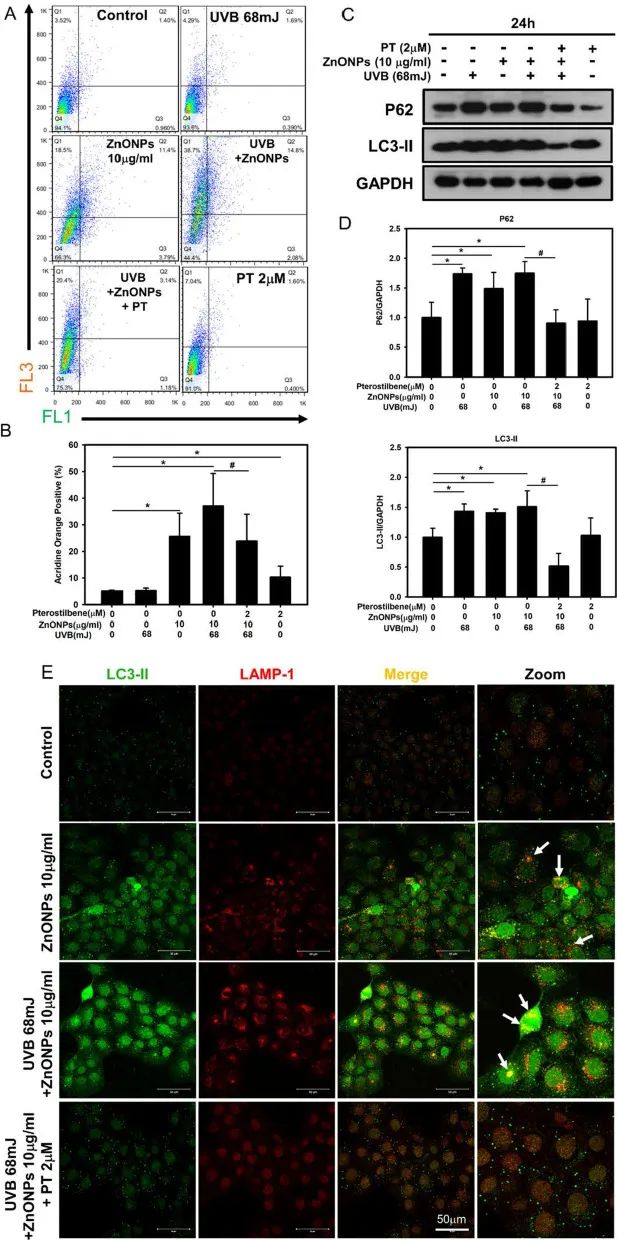
图6. ZnONPs诱导HaCaT自噬功能障碍
6.外泌体介导ZnONPs诱导的NLRP3
在细胞间传递
据报道,细胞内过量的NLRP3炎症小体可以通过外泌体分泌到胞外并影响其他细胞。作者使用超速离心分离出外泌体并检测外泌体中包裹的因子,发现ZnONPs和UVB共同处理后,HaCaT细胞产生的外泌体中包裹有大量的NLRP3及焦亡因子(图7A-E)。当用ZnONPs和UVB共同处理后HaCaT细胞产生的外泌体处理其他细胞时,其NLRP3及焦亡因子显著上调,暗示外泌体将相关蛋白传递至该细胞中(图7F-G)。以上结果表明,ZnONPs和UVB共同处理诱导角质形成细胞产生的NLRP3及焦亡因子可通过外泌体在细胞间传递。
7.自噬功能障碍促进HaCaT细胞
外泌体释放
自噬是外泌体生物发生的关键调节因子之一,自噬体与外泌体前体融合后,可与溶酶体融合降解其内容物。作者使用自噬诱导剂雷帕霉素(Rapa)和自噬抑制剂氯喹(CQ)分别处理细胞,当Rapa处理时由ZnONPs和UVB共同处理后HaCaT细胞外泌体中NLRP3及焦亡因子降低,而CQ处理后则相反(图7H-I)。此外,作者利用Transwell实验构建共培养体系,用Dioc18标记上层HaCaT细胞的外泌体,发现ZnONPs和UVB共同处理上层HaCaT细胞后,其分泌并迁移到下层HaCaT细胞中的外泌体显著增多,Rapa和CQ能分别抑制及促进这一现象(图7J-M)。以上结果表明,ZnONPs和UVB共同处理诱导的角质形成细胞自噬功能障碍会促进其外泌体释放的增强,从而影响其他细胞。
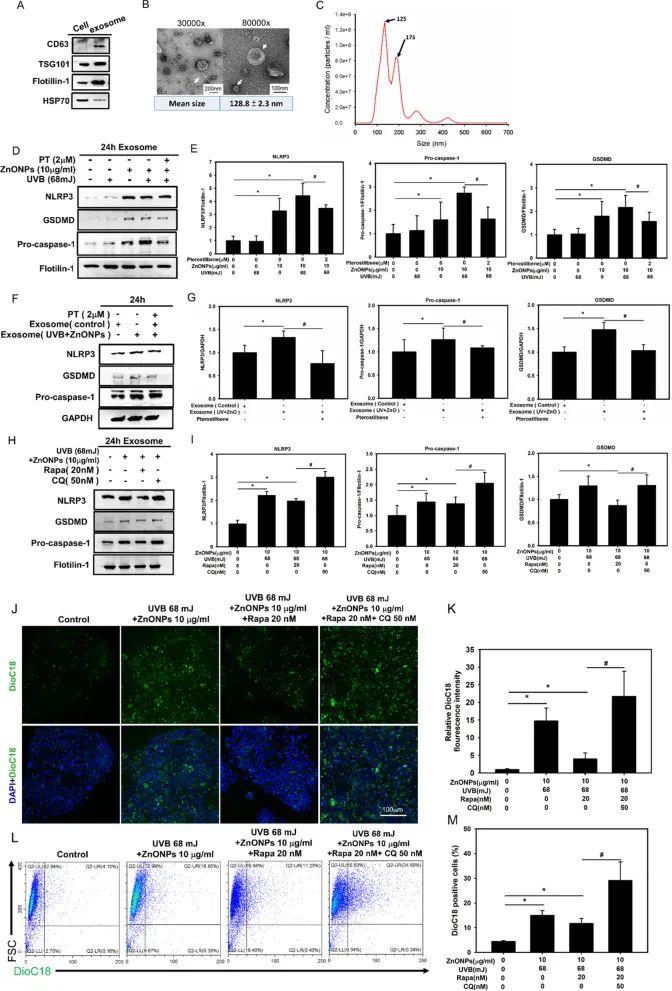
图7. HaCaT细胞释放包含有NLRP3和焦亡因子的外泌体
影响其他细胞
8.PT抑制ZnONPs和UVB对小鼠皮肤的影响
作者使用ZnONPs和UVB共同处理SKH-1无毛小鼠的背部皮肤,发现小鼠背部皮肤明显增厚且经皮水分流失(TEWL)显著降低,PT处理组小鼠则没有出现这种现象(图8A-C)。对小鼠体内的NLRP3炎症小体及焦亡因子检测也发现,ZnONPs和UVB共同处理后炎症及焦亡因子表达量上调,皮肤中发生了炎症及细胞焦亡,而PT的处理则能够下调这些因子的表达量,达到保护皮肤免受损伤的效果。
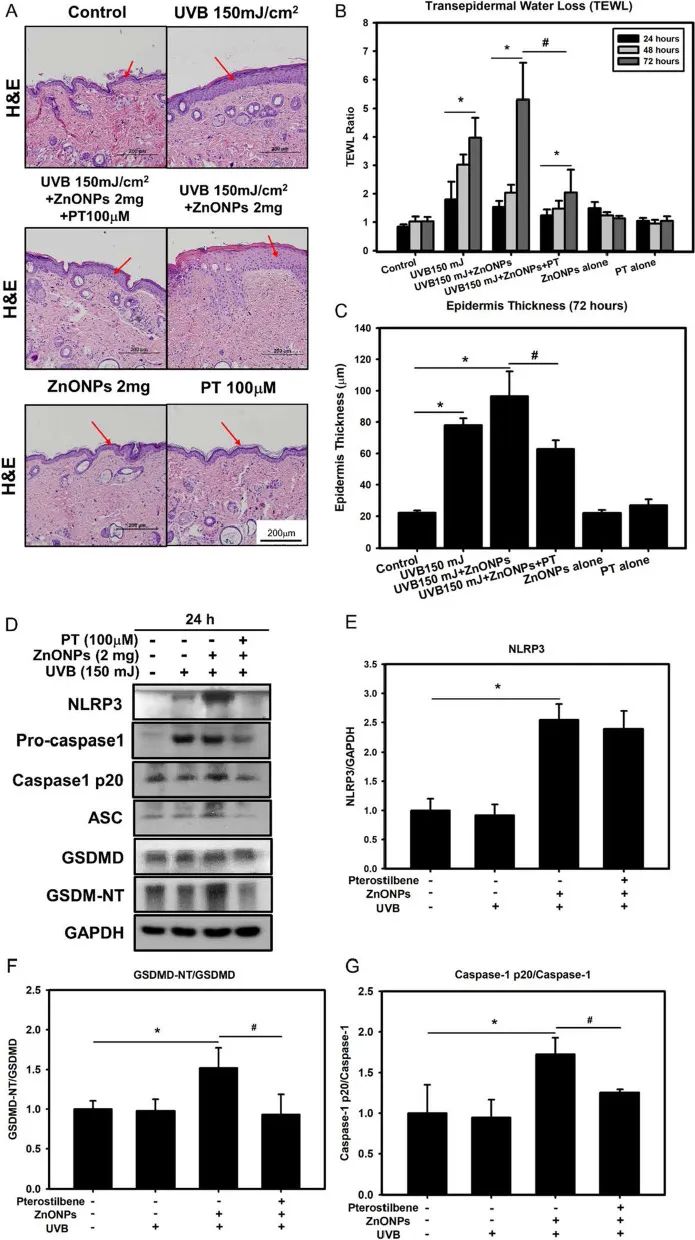
图8. PT保护小鼠皮肤免受ZnONPs造成的损伤
五、总结
在本研究中,作者发现ZnONPs和UVB共同处理HaCaT细胞,一方面会导致细胞内活性氧水平升高以及线粒体破坏从而激活NLRP3炎症因子和焦亡相关因子,引起炎症反应和细胞焦亡;另一方面,ZnONPs会引起HaCaT细胞的自噬功能障碍,使自噬体无法与溶酶体融合形成自噬溶酶体而降解NLRP3炎症因子,使得NLRP3炎症因子及焦亡因子被包裹到外泌体中分泌到细胞外,从而影响其他细胞,造成炎症反应的扩大和皮肤损伤。
该研究对ZnONPs的毒性机制进行了深入的探讨,有助于制定相关纳米材料应用中引起不良反应的预防或治疗措施。同时,文章也对炎症小体、自噬及外泌体之间的关系作出了深入研究,为相关科研热点的研究提供了新的见解。

2022年度国自然医学部国自然40大科研热点的中标数统计如下:
2022热点 | 2022中标数 | 2022热点 | 2022中标数 |
免疫调控 | 907 | 中性粒细胞 | 112 |
巨噬细胞 | 591 | 反馈回路 | 104 |
线粒体 | 491 | 乳酸化 | 104 |
血管功能 | 487 | 可变剪接 | 71 |
外泌体 | 470 | AI机器学习 | 67 |
自噬 | 404 | 类器官 | 67 |
铁死亡 | 337 | 炎症小体 | 62 |
干细胞 | 329 | 染色质重塑 | 58 |
代谢重编程 | 325 | 单细胞测序 | 54 |
m6A/m5C/m7G | 320 | 糖基化 | 50 |
泛素化 | 225 | 低氧缺氧 | 50 |
circRNA | 221 | 相分离 | 50 |
lncRNA | 204 | 泛凋亡PANoptosis | 42 |
细胞焦亡 | 175 | 细胞衰老 | 37 |
组蛋白 | 171 | 胞葬 | 33 |
肠道菌群 | 133 | CRISPR | 33 |
乙酰化 | 125 | 增强子 | 29 |
内质网 | 125 | 精氨酸甲基化 | 25 |
转录调控 | 112 | 迁移体 | 8 |
糖酵解 | 112 | 血管拟态 | 8 |
版权声明:本网站所有注明来源“医微客”的文字、图片和音视频资料,版权均属于医微客所有,非经授权,任何媒体、网站或个人不得转载,授权转载时须注明来源:”医微客”。本网所有转载文章系出于传递更多信息之目的,且明确注明来源和作者,转载仅作观点分享,版权归原作者所有。不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。 本站拥有对此声明的最终解释权。




































 关注公众号
关注公众号 安卓客户端
安卓客户端










发表评论
注册或登后即可发表评论
登录注册
全部评论(0)