
2022-11-21 来源 : 学术查
研究背景
翻译后组蛋白修饰是表观遗传调控机制层次结构的一部分。近年来发现了多种来源于细胞代谢物的组蛋白酰化标记,如丙酰化、丁酰化、2-羟基异丁酰化、琥珀酰化、丙二酰化、戊二酰化、巴豆酰化和β-羟基丁酰化。 乳酸化修饰 (lactylation) 为芝加哥大学赵英明教授团队于2019年在Nature杂志首次报道的一种组蛋白翻译后修饰,糖酵解衍生的乳酸被鉴定为组蛋白乳酸化的底物,并且组蛋白赖氨酸 (K) 残基的乳酸衍生乳酸化 (la) 已被确定为直接刺激染色质基因转录的表观遗传修饰。后续研究进一步证实,组蛋白乳酸化可调节巨噬细胞 M1/2 极化、体细胞重编程和肿瘤发生。然而,目前有关乳酸化修饰的研究尚处于初期阶段,其对于更多疾病和生命活动的具体调控机制尚待进一步发掘。
肿瘤微环境(TME)是一个复杂的生态系统,包含具有促肿瘤和抗肿瘤作用的适应性和先天免疫细胞,显著影响免疫疗法的结果。肿瘤浸润髓系细胞 (TIM) 是参与肿瘤免疫逃逸的关键细胞群,其功能受多种表观遗传机制的调节。RNA N6-甲基腺苷 (m6A) 修饰是 mRNA 中最丰富的修饰,主要由甲基转移酶复合体 (MTC) 进行催化,而METTL3 主要作为复合物中的催化核心发挥重要作用。越来越多的证据表明,m6A 修饰在调节多种细胞功能中起着关键作用。同时,小分子抑制剂靶向 m6A 修饰剂(例如靶向 METTL3和 FTO)在急性髓性白血病(AML)中表现出强烈的抗肿瘤反应,这些发现提出了关于 TIM 中的m6A甲基化是否调节免疫抑制表型的问题,然而m6A 修饰在控制TIM功能中的精确调控模式仍然知之甚少。

研究成果

2022年3月22日,浙江大学基础医学院/良渚实验室王青青教授、来利华副教授与浙大医学院附属二院丁克峰教授团队合作于Molecular Cell杂志(IF 19.328)在线发表“Lactylation-driven METTL3-mediated RNA m6A modification promotes immunosuppression of tumor-infiltrating myeloid cells”研究论文,揭示肿瘤微环境中积累的乳酸通过诱导组蛋白乳酸化修饰上调肿瘤浸润髓系细胞(TIMs)中RNA甲基转移酶 METTL3 表达,提高其 m6A 修饰水平,促进其免疫抑制功能,进而导致肿瘤免疫逃逸。

原文链接:
https://www.sciencedirect.com/science/article/abs/pii/S1097276522002076?via%3Dihub
研究团队对结肠癌患者的组织样本进行分析发现,TIMs 中METTL3的表达上调与结肠癌患者的不良预后相关。动物实验发现,METTL3 的髓系缺乏会显著减弱小鼠的肿瘤生长。通过RNA-seq、m6A 甲基化免疫共沉淀测序等实验手段进行分析,发现METTL3 介导 TIM 中 Jak1 mRNA 的 m6A 修饰,m6A-YTHDF1 轴增强 JAK1 蛋白翻译效率和随后的 STAT3 磷酸化。随后进一步探究METTL3的表达升高的上游机制,发现肿瘤微环境中积累的乳酸通过 H3K18 乳酸化有效诱导 TIM 中的 METTL3 上调,并且课题组在 METTL3 的锌指结构域中发现了两个乳酸化修饰位点,这能够帮助 METTL3 捕获目标 RNA并催化 m6A 修饰的发生。该研究揭示了乳酸化修饰-METTL3-JAK1-STAT3 调节轴能够有效诱导 TIM 的免疫抑制功能,强调了乳酸化驱动的 METTL3 介导的 RNA m6A 修饰对于促进 TIM 的免疫抑制能力的重要性,并提出了METTL3 抑制剂有潜力成为结直肠癌 (CRC) 的新型免疫治疗策略。
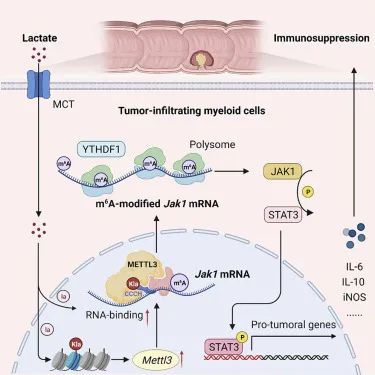
组蛋白乳酸化促进肿瘤免疫逃逸
肿瘤微环境中的乳酸是诱导RNA甲基转移酶METTL3表达和功能的关键因素。METTL3 介导的m6A 修饰有效增强肿瘤浸润性骨髓细胞的免疫抑制功能,促进肿瘤免疫逃逸
研究亮点
1、H3K18 乳酸化增加肿瘤浸润髓系细胞中 METTL3 的表达
2、METTL3 介导的对 Jak1 mRNA 的 m6A 修饰促进其蛋白质翻译
3、METTL3/m6A/JAK1/STAT3轴增强髓系细胞的免疫抑制功能
4、METTL3 锌指结构域的乳酸化增强了其对 m6A修饰 RNA 的捕获
5、METTL3 抑制剂或可成为结直肠癌的新疗法
2022年度国自然医学部国自然40大科研热点的中标数统计如下:
2022热点 | 2022中标数 | 2022热点 | 2022中标数 |
免疫调控 | 907 | 中性粒细胞 | 112 |
巨噬细胞 | 591 | 反馈回路 | 104 |
线粒体 | 491 | 乳酸化 | 104 |
血管功能 | 487 | 可变剪接 | 71 |
外泌体 | 470 | AI机器学习 | 67 |
自噬 | 404 | 类器官 | 67 |
铁死亡 | 337 | 炎症小体 | 62 |
干细胞 | 329 | 染色质重塑 | 58 |
代谢重编程 | 325 | 单细胞测序 | 54 |
m6A/m5C/m7G | 320 | 糖基化 | 50 |
泛素化 | 225 | 低氧缺氧 | 50 |
circRNA | 221 | 相分离 | 50 |
lncRNA | 204 | 泛凋亡PANoptosis | 42 |
细胞焦亡 | 175 | 细胞衰老 | 37 |
组蛋白 | 171 | 胞葬 | 33 |
肠道菌群 | 133 | CRISPR | 33 |
乙酰化 | 125 | 增强子 | 29 |
内质网 | 125 | 精氨酸甲基化 | 25 |
转录调控 | 112 | 迁移体 | 8 |
糖酵解 | 112 | 血管拟态 | 8 |
版权声明:本网站所有注明来源“医微客”的文字、图片和音视频资料,版权均属于医微客所有,非经授权,任何媒体、网站或个人不得转载,授权转载时须注明来源:”医微客”。本网所有转载文章系出于传递更多信息之目的,且明确注明来源和作者,转载仅作观点分享,版权归原作者所有。不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。 本站拥有对此声明的最终解释权。


































 关注公众号
关注公众号 安卓客户端
安卓客户端










发表评论
注册或登后即可发表评论
登录注册
全部评论(0)