2022-08-03

摘要
脓毒症是一种潜在的致命疾病,由机体对感染的免疫反应失调引起。脓毒症引起的肝损伤被认为是重症监护病房死亡的强有力的独立预测因素,有解剖学和流行病学证据表明肠道和肝脏之间存在密切的相互作用。脓毒症期间的肠道屏障破坏和肠道微生物群失调导致肠道病原体相关分子模式和损伤相关分子模式移位到肝脏和体循环中。肝脏通过细菌清除、脂多糖解毒、细胞因子和急性期蛋白释放以及炎症代谢调节等机制对调节全身感染期间的免疫防御至关重要。当肝脏发生不适当的免疫反应或过度炎症时,病原体清除能力受损和肝脏代谢紊乱可导致肠道屏障进一步受损,并增加肠道微生物群组成和多样性的破坏。因此,肠道和肝脏之间相互作用是潜在的治疗靶点。本文综述脓毒症中密切的肠道-肝脏相互作用(肠道-肝脏轴)。
介绍
脓毒症是一种危及生命的器官功能障碍,由宿主对感染的反应失调引起,在全球范围内具有高发病率和死亡率,事实上,在高收入国家,每年有 280 万人死于脓毒症。脓毒症可进展为多器官功能障碍综合征 (MODS) 。MODS 患者的肝功能障碍通常与显著的发病率相关,但其确切患病率仍然未知。肠道微生物群被认为是疾病相关发病率和进展有力指标。例如,毛螺菌科有助于保护肠黏膜屏障,并在脓毒症小鼠模型中提供微生物介导的生存优势。双歧杆菌参与保护大鼠免受脓毒症相关肝损伤。肠道微生物群是免疫系统发育和调节的关键,影响宿主易感性和对感染的反应。肠道菌群失调和肠道通透性增加会促进病原微生物过度生长和肠道病原体相关分子模式 (PAMP) 易位到淋巴和门静脉系统,从而削弱身体对感染或损伤的防御能力,并加重器官损伤。肠道和肝脏之间相互作用得到广泛认可,因为肠道和肝脏通过胆道、门静脉和体循环进行双向交流。此外,肠黏膜和血管屏障充当肠道和肝脏之间的通讯枢纽。肝脏对于改变宿主防御和控制感染中的炎症至关重要。然而,在病理情况下,微生物产物的失调易位引发炎症,损害肝细胞的功能和解毒能力。一般而言,肝功能障碍、细菌清除率降低和代谢紊乱会导致肠道菌群失调加剧和肠道屏障进一步破坏,从而导致 MODS。因此,提高对脓毒症引起的肝损伤期间肠-肝相互作用的了解可能有助于阐明这些复杂的疾病,并为针对潜在机制的脓毒症新治疗方法提供见解。
肠屏障
物理屏障
肠上皮细胞 (IEC) 和粘液成分的存在是维持肠屏障的第一道防线。粘液下方的上皮和肠血管屏障由单层细胞组成,该单层上皮细胞作为保护屏障,限制肠道腔内的病原体、毒素和抗原进入肠系膜淋巴和循环。虽然上皮单层作为主要的物理屏障,但细胞旁间隙受几个细胞间连接的调节,包括顶端紧密连接 (TJ)、下粘附连接和桥粒。值得注意的是,TJ 结构在提供物理屏障以防止腔内炎症分子进入循环中至关重要。粘液覆盖整个肠道表面,由杯状细胞衍生的粘蛋白 (MUC) 组成。小肠被单一的粘液层覆盖,而大肠包含两个主要的粘液层,内部致密的粘液层主要作为肠道的保护屏障,因为它对管腔细菌不渗透。粘液为共生细菌提供碳水化合物,抑制上皮细胞凋亡,并通过充当抗菌肽和免疫球蛋白的粘性陷阱来增加免疫细胞成分的释放。MUC2 是肠粘液层的关键元素。小鼠缺乏 Muc2 会加剧亚致死剂量的脂多糖 (LPS) 诱导的肠道细菌易位至肝脏并增加死亡率。此外,肠上皮产生大量抗菌肽 (AMP) 和肠碱性磷酸酶 (IAP) 。AMP 通过分泌溶菌酶、调节下游先天免疫反应和干扰细菌细胞壁合成等来快速杀死或灭活细菌,这是一种随着时间的推移而进化的免疫防御方法。IAPs可以防止新释放的LPS的后续毒性,调节HCO3-分泌,促进肠道共生菌的生长。值得注意的是,IAP 还阻止 LPS 触发 toll 样受体 (TLR)-4/髓样分化因子 88 (MyD88) 介导的炎症级联反应。最近的研究表明,IAP 诱导 IEC 和巨噬细胞中的自噬,在小鼠中发挥抗炎作用。
免疫屏障
肠道中的免疫细胞形成第二道防线以维持肠道屏障(表1)。在解剖学上,肠管壁从内到外分为黏膜、黏膜下层、肌层和浆膜层四层。丰富的淋巴滤泡存在于黏膜和黏膜下组织的固有层中,包括分离和聚集的淋巴滤泡。考虑到它们在肠壁的分布,肠道免疫细胞主要包括肠淋巴滤泡、上皮间淋巴细胞和黏膜固有层中的淋巴细胞。肠上皮屏障受到局部上皮间淋巴细胞的保护,这些淋巴细胞迅速激活针对受感染或受压上皮的 T 辅助 1 (Th1) 细胞细胞因子反应。有趣的是,体外插入肠道上皮细胞的树突状细胞(DCs)可以打开IECs之间的TJs并直接摄取管腔微生物。此外,DC 表达 TJ 蛋白以保持上皮屏障的完整性。黏膜固有层包括浆细胞样 DC、先天淋巴细胞、黏膜相关不变 T 细胞和攻击病原体的 T 细胞。分泌型免疫球蛋白 A (IgA) 在由固有层浆细胞介导的适应性免疫中发挥关键作用:它调节肠道微生物群组成,保护肠道上皮免受病原微生物的侵害,并有助于免疫系统发育。通过将病原体抗原呈递给肠道免疫细胞,IEC 有助于肠道适应性免疫。膜细胞是特化的上皮抗原呈递细胞,分散在与卵泡相关的上皮细胞中,对抗原特异性 IgA 的产生至关重要。这些与肠道相关的免疫机制可保护全身循环免受肠道病原体的有害影响。
肠道共生菌群
人类微生物是多种多样的,估计有 100 万亿微生物由 500 到 1000 种不同的细菌物种组成。大多数已被证实由五个门组成,主要是厚壁菌门(79.4%)和拟杆菌门(16.9%)。通常,共生微生物群会影响肠道环境,限制侵入性病原体的生长。例如,微生物衍生的代谢物在体外抑制大肠杆菌O157 的生长。共生细菌还产生主要由厚壁菌门产生的细菌素,以对抗入侵的病原体。其他细菌,包括变形菌门、拟杆菌门和放线菌门,也编码各种细菌素。此外,被称为益生菌的几种产生细菌素的共生细菌(双歧杆菌和乳酸杆菌)可用于促进肠道健康。整个胃肠道微生物群落的分布、适应性和功能与不同的好客环境条件相匹配,使宿主和共生微生物群之间互惠互利。健康成人的胃中细菌最少,即乳酸杆菌和螺杆菌,而十二指肠微生物群以厚壁菌门和放线菌为主。厚壁菌门在回肠和结肠中继续占主导地位(43%),但在回肠和结肠中变形菌和拟杆菌的丰度稳步上升。与小肠相比,大肠显示出更多的微生物种类,尽管结肠往往被两个重要的门占据(拟杆菌门占 50%,厚壁菌门占 45%)。肠道微生物通过加强肠道屏障来促进健康,共生细菌通过竞争营养和空间来抵御病原体。例如,共生肠杆菌科通过在小鼠体内竞争氧气来对抗沙门氏菌定植。此外,共生细菌激活宿主模式识别受体以增强 AMP 和 MUC 的产生。它们还通过提供适度水平的免疫刺激来诱导 IgA 分泌,因此建立了一种基本的免疫适应,在使宿主和微生物群落处于稳态中发挥着至关重要的作用。肠道微生物群产生的短链脂肪酸可作为共生细菌的能量来源,并防止 LPS 引起的肠道屏障紊乱。总体而言,微生物群失调被认为是各种疾病的主要诱因。
肠-肝轴
生理“肠-肝轴”
肝脏是最大的腺体,分泌最终排入小肠的胆汁酸,并由混合各种肝脏结构和细胞的动静脉血提供。富含氧气的动脉血通过肝动脉进入肝脏,但它是该器官的次要血液供应。相反,进入肝脏的门静脉具有丰富的营养物质和 LPS 等病原体衍生分子,是肝脏的主要血液供应。正常情况下,肝脏通过胆道系统和体循环与肠道及其微生物群进行交流(图 1 )。肠道及其微生物群与肝脏的双向关联被称为生理“肠-肝轴”。
在脓毒症期间,多种机制导致肠道屏障的破坏,包括 IEC 凋亡、粘液层的改变和细胞间连接的破坏,导致肠道 PAMP 通过淋巴管、门静脉循环或胆道易位进入肝脏。肝脏对于免疫防御的调节至关重要,LSEC、巨噬细胞、星状细胞和肝细胞等效应细胞可立即识别和参与病原体、清除细菌和释放细胞因子。当肝脏中出现高水平的 DAMP 和促炎细胞因子产生不适当的免疫反应或过度炎症时,肝窦的正常结构被破坏,这些细胞通过凋亡和自噬而受损,导致细菌清除功能障碍和代谢紊乱。结果,肠道屏障进一步受损,肠道菌群失调加剧,远端器官因PAMPs和DAMPs的扩散和全身炎症而受损。IECs,肠上皮细胞;DAMP,与损伤相关的分子模式;LSECs,肝窦内皮;PAMPs,病原体相关分子模式
肝脏的免疫防御
肝脏也是关键的免疫组织,肝窦内皮细胞 (LSEC)、巨噬细胞、星状细胞、淋巴细胞和胆管细胞构成了非实质细胞的大部分。LSECs构成肝窦血管或毛细血管床的内层,富含营养的肝门静脉血液和富含氧气的肝动脉血液混合,通过检测和捕获病原体,甚至可能呈递抗原和去除大分子废物,有助于免疫监视来自血液。枯否细胞 (KCs) 构成健康肝脏中的大部分肝巨噬细胞。源自卵黄囊的集落刺激因子 1 受体阳性红髓祖细胞和源自骨髓的单核细胞都是 KCs 的来源。人类肝脏包含两种不同的巨噬细胞亚型:CD68 +具有胶原结构的巨噬细胞受体 (MARCO) +巨噬细胞,有助于维持免疫耐受和炎症抑制,以及 CD68 + MARCO-巨噬细胞,它们更具促炎性。
脓毒症对肠道屏障和微生物群的影响
炎症显著促成了脓毒症中肠道损伤的发病机制 。脓毒症影响 claudins、连接粘附分子 A、occludin 和zonula occludens-1的表达,并激活肌球蛋白轻链激酶,调节肠道通透性。由于PAMP诱导的肠干细胞上表达的 TLR4 被激活,如 LPS,脓毒症可能直接影响肠干细胞的生长和凋亡。肠道高通透性与失调的 IEC 细胞凋亡密切相关,导致粘液层发生变化,例如厚度减少、管腔覆盖率降低和附着力差。同样,绒毛长度减少与肠道通透性增加和 IEC 细胞凋亡有关。Bcl-2过表达通过在脓毒症后发生高渗透性的转基因小鼠中的 TJ 改变来提高存活率。肠道生理机能紊乱之后是脓毒症患者的微生物群失调。在重症脓毒症患者中,肠道微生物群的多样性下降,其组成以多重耐药菌为主。广谱抗生素治疗可以显著改变重症患者的肠道微生物群。例如,氟喹诺酮类药物增加了乳球菌和片球菌,但减少了重症患者的大肠杆菌/志贺氏菌和脱硫弧菌。
脓毒症中的肠-肝轴
脂多糖的具体作用
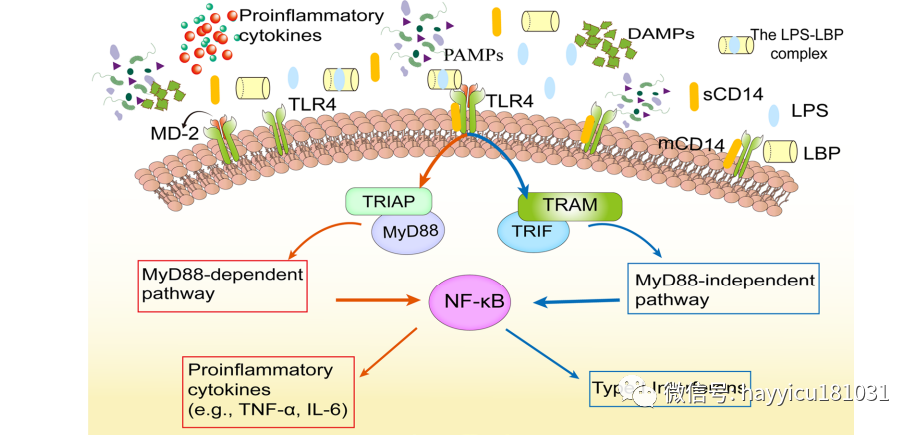
人体肠道是来自估计 100 万亿微生物的 ≥ 1 g LPS 的储存库,即使在健康的人体血浆中也可以检测到内毒素。根据其大小,LPS 分子在穿过小肠时通过多个替代路径,包括细胞旁通路、网格蛋白介导的内吞作用、小胞饮作用和脂筏介导的内吞作用、杯状细胞相关抗原通道和乳糜微粒途径。在结肠中,LPS 可以通过网格蛋白介导或囊泡介导的蛋白质转运途径转运。当肠道屏障的成分受到脓毒症炎症的影响时,如 IEC 细胞凋亡失调所指出的,这些有助于防止大量 LPS 转移到体循环的防御腔机制逐渐失效,导致 LPS 迁移到多个器官和触发不受控制的免疫炎症反应。通常,通过门静脉的肠道 LPS 由肝脏处理和解毒。LPS 的解毒是由清道夫受体 (SR) 介导的肝脏摄取所吞噬,由酰基水解酶 (AOAH) 和碱性磷酸酶进行。AOAH 选择性地去除脂质 A 级分中与主链相连的二级脂肪酰基链,在控制 LPS 毒性方面发挥重要作用。此外,血浆中含有具有不同亲和力的 LPS 结合蛋白,包括 LPS 结合蛋白 (LBP)、CD14、杀菌/通透性增加蛋白 (BPI) 和参与 LPS 解毒的脂蛋白。LBP 主要由小鼠脓毒症急性期的 IEC 和肝细胞产生。它与 LPS 的脂质 A 特异性结合,形成 LPS-LBP 复合物;这有助于 LPS 转移并与单核细胞/巨噬细胞和中性粒细胞表面的膜 CD14 结合,最终被 TLR4 和骨髓分化-2 识别,促进炎症反应级联反应。TLR4 信号通路的激活,分类为 MyD88 依赖性、MyD88 非依赖性和含有 Toll/IL-1R 结构域的适配器诱导干扰素 (IFN)-β 依赖性途径,导致细胞因子和细胞损伤显著增加(图 2 )。然而,LBP 和可溶性 CD14 的高血浆浓度可能有助于限制对 LPS 的有害全身反应。BPI 在肠上皮细胞中表达,受损细胞中钾水平的变化可作为损伤相关分子模式 (DAMP) 促进小鼠肠上皮细胞中 BPI 的表达。LPS激活的TLR4信号通路概述。LBP 和 CD14 促进的 LPS 识别由 TLR4 和 MD-2 受体复合物介导。TLR4 信号通路的激活分为 MyD88 依赖性、MyD88 非依赖性和 TRIF 依赖性通路,介导促炎细胞因子(TNF-α、IL-6 等)和 I 型干扰素基因的激活。IL,白细胞介素;LBP,LPS结合蛋白;LPS,脂多糖;MD-2,骨髓分化-2;MyD88,骨髓分化因子 88;TLR4,toll样受体 4;TRIF,含 Toll/IL-1R 结构域的适配器诱导 IFN-β;TNF,肿瘤坏死因子。
肠道功能障碍对肝脏的影响
脓毒症伴随肠道功能障碍后,肠道中的 PAMP 和 DAMP 通过门静脉循环或胆道到达肝脏。在肝脉管系统中,效应细胞立即参与循环病原体或识别 PAMP,形成免疫哨兵。此外,肝脏中巨噬细胞和其他细胞类型上的 TLR 被来自垂死宿主细胞的内源性成分(称为 DAMP)激活。NLR 和 RNA 解旋酶家族也能识别细胞质中的病原体。肠道衍生的 PAMP 和 DAMP 可能是关键触发因素,导致不适当的免疫反应或严重炎症、肝脏病原菌清除受损和代谢紊乱(表2)。
肝窦内皮细胞
由于 LSEC 在内皮下方缺乏基底层或隔膜,因此它们在窦状血液和内皮下空间之间提供称为窗孔的通信区域,促进血液与附近星状细胞和肝细胞之间的底物交换并调节后者中的脂蛋白交通。血压和毒素水平会影响小鼠 LSEC 窗孔的直径。LSEC 免疫球蛋白超家族的粘附分子在白细胞迁移到炎症部位中起关键作用。正常 LSEC 非常微弱地表达血管细胞粘附分子 1,但在大鼠炎症刺激下其表达显著增加。LSEC 具有极高的内吞能力,由 SR 和溶酶体活性促进,这有助于几种废物和小胶体颗粒的内化和分解代谢。例如,SR-A 1/1.1 型与小鼠体内的各种大分子结合,例如 LPS 和脂磷壁酸 (LTA) 。目前正在 LSEC 中研究 7 个 TLR。在鼠 LSEC 上表达的 TLR-9 识别并结合细菌 DNA 中丰富的未甲基化 CpG 基序,以激活核因子 kappa-B (NF-κB) 并刺激 IL-1β 和 IL-6 的产生。此外,LSEC 通过 TLR3 启动抗病毒和促炎反应,并通过 TLR1、TLR6 和 TLR8 启动适应性反应。因此,LSEC 在肝脏中形成解剖筛,持续监测具有适当耐受性PAMP 和 DAMP等抗原,维持肝脏免疫稳态。
巨噬细胞
巨噬细胞表现出可变的表型,包括通常由 TLR 配体和 IFN-γ 触发的 M1 样巨噬细胞和由 IL-4/IL-13 激活的 M2 样巨噬细胞,以及介于之间的各种状态。促炎细胞因子由 M1 表型显著产生,它还产生活性氮和氧中间体,促进 Th1 免疫反应,并在杀微生物和杀肿瘤活性中发挥重要作用。Th2 偏向反应与 M2 表型有关,这通常与 IL-10 和 IL-1 受体拮抗剂的产生有关。当巨噬细胞暴露于脓毒症小鼠肠道中的细菌产物时,它们会极化为 M1 表型并参与免疫炎症反应。M2 样巨噬细胞可减轻脓毒症引起的大鼠急性肝损伤,可能是通过上调 IL-10 表达和抑制 TNF 产生。增强 M2 样巨噬细胞极化和抑制 M1 样巨噬细胞极化均可保护肝脏免受过度炎症诱导的损伤(图 3)。Neochromine S5 可以改善小鼠的致死性脓毒症,特别是由于其对 M1 样巨噬细胞的调节作用。当肠源性PAMPs暴露于肝巨噬细胞时,巨噬细胞发生极化,形成大量M1样巨噬细胞,主要产生IL-1β、TNF、IL-6等促炎细胞因子;一些 M2 样巨噬细胞通常会产生 IL-10 并在抗炎反应中发挥作用。炎症小体在肝巨噬细胞中被激活,并响应病原体感染和组织损伤。此外,中性粒细胞被趋化因子(如源自 KCs 的 CXCL1 和 CXCL2)吸引到肝脏,释放的 NETs 参与清除病原体和毒素。血小板募集对于限制细菌感染也很关键,与 KCs 相互作用的血小板在对抗细菌感染方面起着至关重要的作用。然而,当肝脏中产生高水平的 DAMP 和促炎细胞因子产生不适当的免疫反应或严重炎症时,就会发生显著的肝细胞损伤、巨噬细胞自噬和细胞凋亡。此外,KCs 在防御病原体方面还有一些特殊的作用。一方面,免疫球蛋白超家族 (CRIg) 的补体受体在各种组织中的常驻巨噬细胞亚群上表达,尤其是在肝脏 KCs 上,它识别补体成分 C3 的活化形式,循环和有效清除调理的传染性病原体。小鼠CRIg基因缺陷显著降低金黄色葡萄球菌和单核细胞增生李斯特菌的 KC 摄取。CRIg 还通过与小鼠体内的 LTA 结合,直接从循环中捕获革兰氏阳性菌。细菌如蜡状芽孢杆菌和耐甲氧西林金黄色葡萄球菌在小鼠体内被 KCs 迅速捕获并触发血小板改变其粘附模式以维持糖蛋白 IIb 介导的 KC 表面粘附以包裹微生物,防止病原体诱导的内皮通透性和肝损伤。然而,在处理巨噬细胞-血小板相互作用时,重要的是要考虑免疫反应和潜在并发症,例如血栓形成。
中性粒细胞、NK 细胞和自然杀伤 T (NKT) 细胞
肝脏微循环中活化中性粒细胞的积聚可能导致免疫介导的损伤。中性粒细胞胞外陷阱 (NET) 在脓毒症期间由中性粒细胞释放,当它们迁移到肝血窦时。LPS 或脓毒症触发由小鼠血窦内 β2-整合素依赖性血小板-中性粒细胞相互作用介导的血管内 NET 发育。此外,中性粒细胞-内皮细胞相互作用和 NETs 在脓毒症大鼠模型中诱导微血栓形成和血管渗漏。NK细胞在肝脏中比在循环中丰富得多;具有几个不同亚群的 NKT 细胞有助于抗菌防御,并受肠道微生物群的影响。NK 和 NKT 细胞都有助于肝损伤的病理生理学。例如,在小鼠暴发性肝炎模型中注射聚 I:C 和 D-半乳糖胺后,NK 细胞通过 NK 组 2 成员 D/配体识别与 KCs 相互作用,导致严重的肝损伤。此外,当肝脏发生缺血或毒素诱导的损伤时,活化的 NKT 细胞主要分泌 IFN-γ,导致中性粒细胞和巨噬细胞的积累,促进小鼠的肝损伤。另一项研究表明,在小鼠模型中,NKT 的激活可以直接通过 Fas/FasL 途径杀死肝细胞。
治疗
上皮屏障靶向治疗
由于 claudin-2 在门控细胞旁通道形成和 TJ 通道调节中的作用,它可能是调节上皮屏障的理想治疗靶点。事实上,调节 TJ 通道门控动力学和蛋白质分子间相互作用可能对炎症相关的屏障失效具有治疗价值,例如 occludin S408 去磷酸化。然而,需要更多的研究来确定为治疗目的调节门控活性的药理学方法。此外,microRNA-155 可减轻脓毒症小鼠的炎症和肠道屏障功能障碍,通过使 NF-κB 信号失活降低 TNF-α 和 IL-6 水平。
针对肠道微生物组
益生菌是活的非致病微生物,有助于保护肠道屏障、抑制病原体传播、最大限度地减少细菌置换和预防感染。然而,流行病学调查中的基因组分析检测到 ICU 患者有六起独立的益生菌从胶囊转移到血液中的事件,从而导致菌血症。益生元是难消化的膳食成分,其中大部分包含难消化低聚糖,可特别促进有益肠道微生物群的发育和代谢,改善肠道菌群的平衡,有益于人类健康。在小鼠脓毒症模型中,纤维素补充剂与部分减少全身炎症有关,这表明微生物赋予了生存益处。粪便微生物移植 (FMT) 可以使小鼠免于由从脓毒症患者身上分离出的病原体引起的致命脓毒症,这与产生丁酸盐的拟杆菌的传播、病原体清除率的提高以及通过干扰素调节因子 3 恢复宿主免疫力有关。FMT 可能是治疗脓毒症的一种选择;然而,供体筛查对于防止可能导致不利感染的细菌传播是必要的。此外,重要的是要考虑 FMT 在不同患者群体中的优势和危害。一种完全不同的策略是使用吸收性材料来防止肠源性毒素和细菌产物进入血液循环和肝脏。事实上,针对高钾血症和高磷血症的阳离子交换树脂已显示出疗效。不可吸收纳米孔碳降低了接受胆管结扎的大鼠的门静脉压力和肝脏生化标志物,从而减少了内毒素诱导的 KC 刺激。因此,通过靶向肠-肝轴,吸附材料是脓毒症的一种可能治疗策略。
高密度脂蛋白 (HDL) 的潜在疗法
一项研究表明,HDL 能够通过 SR-BI 介导的小鼠 LPS 摄取来中和 LPS 并加速 LPS 清除。HDL颗粒上的许多载脂蛋白(Apo)在清除内毒素以预防感染方面起着至关重要的作用。例如,当 LPS 与 Apo AI 或 Apo E 结合时,对 LPS 炎症反应受到抑制,但 Apo AII 和 Apo CI 与 LPS 结合,从而增强炎症。在一项针对 63 名严重脓毒症患者的研究中,第 1 天 HDL 水平 < 20 mg/dL 和 Apo AI < 100 mg/dL 与总体死亡率和可归因于脓毒症的 30 天死亡率、重症监护病房住院时间延长和医院获得性感染。此外,重组 HDL 输注可减少内毒素诱导的大鼠肺、肝和肠损伤。此外,小鼠中 111-铟细菌标记强调HDL 摄取促进潜在的肝脏细菌清除的可能性。因此,HDL 可能是未来脓毒症预防和治疗重要目标。
结论
脓毒症期间,由于炎症导致肠道屏障成分逐渐受损,导致肠道通透性增加,肠道失调,包括病原微生物过度生长,大量PAMPs和DAMPs通过门静脉循环或胆道转运到肝脏。由于肝脏在物理上靠近肠道,作为一线免疫器官,它在肠道和体循环之间扮演着守门人的角色,可以迅速从免疫低反应状态转变为产生强有力的炎症反应和有效的适应性免疫。然而,脓毒症后肠道高通透性和肠道源性PAMPs和DAMPs可引发不适当的免疫反应或压倒性的炎症,导致肝脏清除致病菌受损和代谢紊乱,导致MODS。因此,深入了解脓毒症的肠-肝相互作用,有助于预防和限制脓毒症引起的肝损伤,改善脓毒症患者的预后。
图1在脓毒症期间,几种机制有助于肠道屏障的破坏,包括IEC凋亡、粘液层的改变和细胞间连接的破坏,导致肠道PAMP通过淋巴管、门静脉循环或胆道转移到肝脏。肝脏对免疫防御的调节至关重要,其反射细胞(如LSECs、巨噬细胞、星状细胞和肝细胞)可立即识别和接触病原体,清除细菌,并释放细胞因子。当肝脏中高水平的DAMP和促炎症细胞因子产生时,出现不适当的免疫反应或压倒性炎症反应,肝窦的正常结构被破坏,这些细胞通过凋亡和自噬受损,导致细菌清除功能障碍和代谢紊乱。结果,由于PAMPs、DAMPs和系统信息的传播,肠道屏障进一步受损,肠道菌群失调加剧,远端器官受损。肠上皮细胞;DAMPs,损伤相关分子模式;LSECs,肝窦内皮;PAMPs,病原体相关的分子模式。
图2 LPS激活TLR4的信号通路概述。
由LBP和CD14促进的LPS识别由TLR4和MD-2受体复合物介导。TLR4信号的激活分为MyD88依赖性、MYD1988非依赖性和TRIF依赖性途径,介导促炎症细胞因子(TNF-α、IL-6等)和I型干扰素基因的激活。IL,白细胞介素;LBP,LPS结合蛋白;脂多糖;MD-2,髓样分化-2;MyD88,髓样分化因子88;TLR4,toll样受体4;TRIF,含有适配器的Toll/IL-1R结构域诱导IFN-β;肿瘤坏死因子。
图3当肠源性PAMP暴露于肝巨噬细胞时,巨噬细胞极化并形成大量M1样蛋白,巨噬细胞主要产生促炎细胞因子,如IL-1β、TNF和IL-6;一些M2样巨噬细胞通常产生IL-10,并在抗炎症反应中发挥作用。内质体在肝巨噬细胞中被激活,并对病原体感染和组织损伤作出反应。此外,嗜中性粒细胞被趋化因子吸引到肝脏,例如来自KCs的CXCL1和CXCL2,并且释放的Net参与去除病原体和毒素。血小板聚集对于控制细菌感染也至关重要,与KC相互作用的血小板在抑制细菌感染方面发挥着至关重要的作用。然而,当不适当的免疫反应或压倒性炎症发生时,肝脏中高水平的潮湿形成和促炎症细胞因子产生,就会出现显著的肝细胞损伤、巨噬细胞自噬和凋亡。肝脏巨噬细胞由KC增殖和循环单核细胞募集和分化补充。CXCL,趋化因子(C-X-C基序)配体;DAMPs损伤相关的分子模式;IL,白细胞介素;KCs,Kupfer细胞;中性粒细胞胞外陷阱;PAMPs,病原体相关分子模式;肿瘤坏死因子。
---Crit Care 2022 Jul 13;26(1):213. doi: 10.1186/s13054-022-04090-1.
百度浏览 来源 : 重症沙龙

版权声明:本网站所有注明来源“医微客”的文字、图片和音视频资料,版权均属于医微客所有,非经授权,任何媒体、网站或个人不得转载,授权转载时须注明来源:”医微客”。本网所有转载文章系出于传递更多信息之目的,且明确注明来源和作者,转载仅作观点分享,版权归原作者所有。不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。 本站拥有对此声明的最终解释权。















































 关注公众号
关注公众号 安卓客户端
安卓客户端










发表评论
注册或登后即可发表评论
登录注册
全部评论(0)