
2022-11-22 来源 : 学术查
研究背景 ✦
“铁死亡”概念自提出之日就引起了人们的极大关注。尤其是近些年来,铁死亡已成为抗癌治疗中绕过化疗耐药性和消除转移性肿瘤的有力工具。此外,新开发的铁死亡诱导剂已与基于纳米技术的肿瘤细胞靶向结合,为以后的抗癌治疗提供了一个不错的前景。然而,近期有研究报道肿瘤细胞的铁死亡会抑制免疫细胞的功能,促进肿瘤的免疫逃逸。这不得不引起人们对铁死亡如何影响肿瘤微环境的深思。
研究成果
2022年6月27日,VIB-UGent 炎症研究中心,Peter Vandenabeele研究团队于2022年7月在Nature Communications杂志(IF=17.7)上发表题目为“Cancer cells dying from ferroptosis impede dendritic cell-mediated anti-tumor immunity”的文章。该研究揭示了诱导铁死亡后,产生铁死亡的肿瘤细胞会对周围抗原递呈细胞产生抑制作用。

原文链接:https://www.nature.com/articles/s41467-022-31218-2
铁死亡,指的是一种依赖铁的细胞程序性死亡,其特征是出现细胞内氧化还原平衡被破坏和过度脂质过氧化。最近,铁死亡疗法已经成为一种新兴的抗肿瘤治疗方式,可以避免出现传统的耐药,达到清除已转移肿瘤的目的,目前,铁死亡疗法已经和纳米材料和脂质体等不同技术相融合,在肿瘤治疗领域大放异彩。
然而,尽管铁死亡诱导的治疗应用前景光明,但铁死亡如何与免疫系统相互作用仍不清楚。因此,人们越来越需要了解这种新发现的细胞死亡形式与癌症(尤其是免疫疗法)之间的关系。在过去的十年中,癌细胞的免疫原性细胞死亡(ICD)的概念已经被提出,它是一种细胞死亡的类型,导致针对癌细胞的细胞毒性T细胞(CTL)反应的产生,并有助于肿瘤的根除。诱导免疫原性细胞死亡是肿瘤细胞与免疫系统相互作用的三个组成部分的结果,即:树突状细胞(DC)表面肿瘤相关抗原(TAA)的传递、加工和呈递,传播佐剂性的损伤相关分子模式(DAMP)的释放,趋化因子、细胞因子和干扰素驱动的免疫刺激。
1. 发生铁死亡的肿瘤细胞不能引起免疫反应
预防性癌症疫苗接种模型是测试特定细胞死亡方式的免疫原性的有力方法。为此,作者首先将死亡的肿瘤细胞注射在一侧腹部,然后通过随后在另一侧皮下注射活癌细胞来测试对肿瘤发生的保护(图1a)。作者首先选用了3种铁死亡诱导药物,分别对应早期、中期、晚期的三期铁死亡进展,并通过体内实验证明,当采用ML162药物诱导的铁死亡细胞作为肿瘤疫苗,并不能显著表现出抗肿瘤活性。而后,通过对于ML162、RSL-3、Erastin三种铁死亡诱导剂处理后的细胞上清进行分析,发现其中具有DAMP和CXCL1、TNF和IFN-β等细胞因子的释放(图1b)。而对于钙网蛋白的调节,铁死亡诱导后虽然能够上调钙网蛋白的表达,但是与凋亡相比,铁死亡诱导的钙网蛋白高表达主要出现在一些死细胞的胞膜表面。这些数据表明,尽管存在与免疫原性细胞死亡相关的DAMP和细胞因子的释放和暴露,但铁死亡并不是一种免疫原性细胞死亡类型。
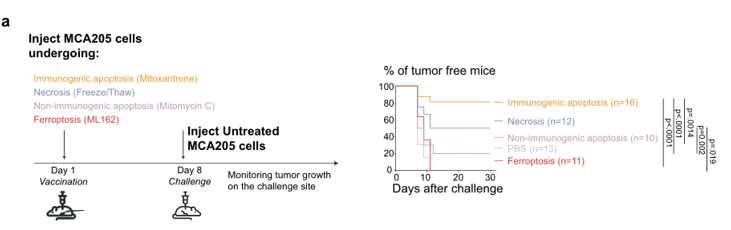
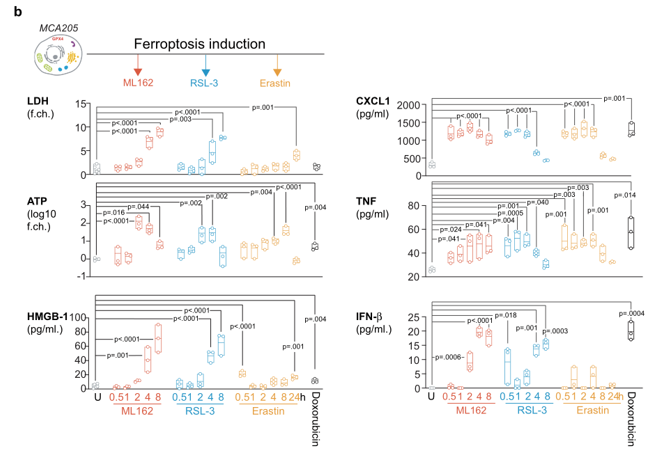
图1 (a)采用MCA205细胞预防接种模型,通过诱导对肿瘤生长的保护作用,评估铁死亡(ML162, 0.5µM, 14 h)的免疫原性潜力。以免疫原性(米托蒽醌,1µM, 24 h)和非免疫原性(丝裂霉素C, 30µM, 24 h)凋亡和意外坏死(冷冻/解冻,3个循环)条件作为对照。Kaplan-Meier曲线表示注射活癌细胞后无瘤小鼠的百分比。n = 3个独立实验数据采用Kaplan-Meier简单生存分析。(b)MCA205细胞被ML162(0.5µM)、RSL-3(0.5µM)、Erastin(2.0µM)和阿霉素(1µM, 24 h)刺激后的DAMP释放情况。分析细胞死亡诱导后不同时间点LDH、ATP、HMGB1以及细胞因子CXCL1、TNF和IFN-β的释放情况。
2. 无论细胞处于铁死亡的哪个阶段,都没有免疫原性。
为了进一步研究铁死亡过程中的DAMP和CXCL1、TNF和IFN-β等细胞因子的释放,并更好地描述铁死亡细胞释放的免疫调节因子的动力学,作者构建了一个多西环素(Doxycycline,Dox)诱导的GPX4敲低细胞模型,用以操控铁死亡进程(图2a,b).DOX诱导后8h,细胞死亡率可至100%。并且通过ROS表达、细胞因子和DAMP分泌及钙网蛋白表达三个层面验证,可以用过调控DOX处理的时间,来操控肿瘤细胞铁死亡进程的时期(图2c)。而后体内实验证明,无论免疫早期还是晚期铁死亡细胞,都无法作为肿瘤疫苗,影响小鼠生存周期(图2g)。
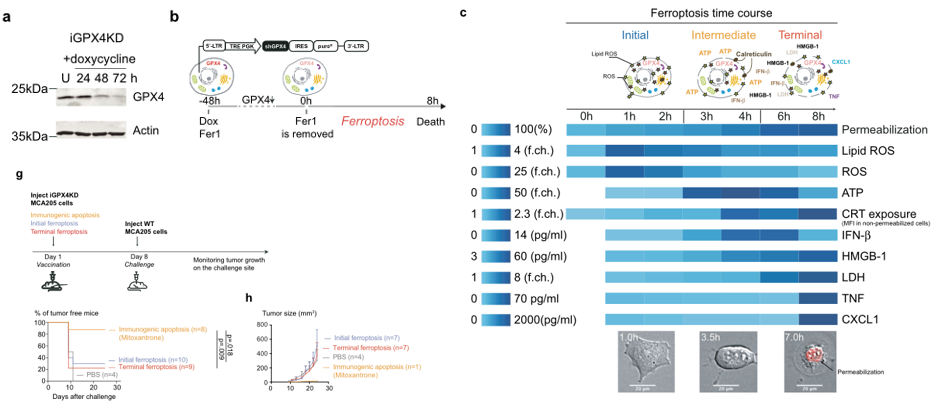
图2(a)多西环素给药(1µg/ ml)诱导iGPX4KD MCA205细胞系GPX4敲除,western blotting检测。(b)诱导iGPX4KD细胞铁死亡的方案。(c)分析脂质ROS、细胞内胞质ROS的积累、死亡细胞、非通透性细胞表面的钙网蛋白水平,以及在铁死亡过程中细胞释放的ATP、HMGB1、LDH、IFN-β、tnf-α和CXCL。铁死亡可分为三个阶段:最初的铁死亡,细胞经历脂质ROS的积累;涉及部分渗透作用和ATP释放以及钙网蛋白暴露的中期铁死亡,以及涉及完全渗透作用和LDH、HMGB1和细胞因子释放的晚期铁死亡。(g)细胞死亡初期和终末期iGPX4KD细胞预防接种模型。米托蒽醌处理(1 μM, 24 h)的免疫原性凋亡的iGPX4KD细胞作为阳性对照。
3. 初期的铁死亡损害树突状细胞的成熟
为了阐明铁死亡的癌细胞为何不是免疫原性的(尽管释放了DAMP、细胞因子、趋化因子和IFNγ),作者进行了一系列实验来研究铁死亡癌细胞与DC之间的相互作用。作者使用ML162、RSL-3、Erastin诱导肿瘤细胞铁死亡后,与DC细胞进行共孵育,发现均可以提高DC细胞的成熟度。为了细化实验步骤,他又用iGPX4的细胞模型进行双重验证(图3a),发现在肿瘤细胞发生铁死亡早期,不仅不能促进DC细胞成熟,反而会抑制DC细胞的活力。另外暴露了一个很有意思的现象,ML162诱导的早期铁死亡,能够抑制DC细胞对于细胞因子的释放,这是铁死亡细胞明显区别于UV诱导细胞细胞凋亡的区别(图3c)。除此以外,铁死亡区别于凋亡,当细胞存活时,磷脂酰丝氨酸并不会外翻,影响DC细胞对于肿瘤细胞的识别。而DC细胞与早期铁死亡细胞共培养后,也会导致DC细胞脂滴堆积,影响DC细胞功能(图3h)。
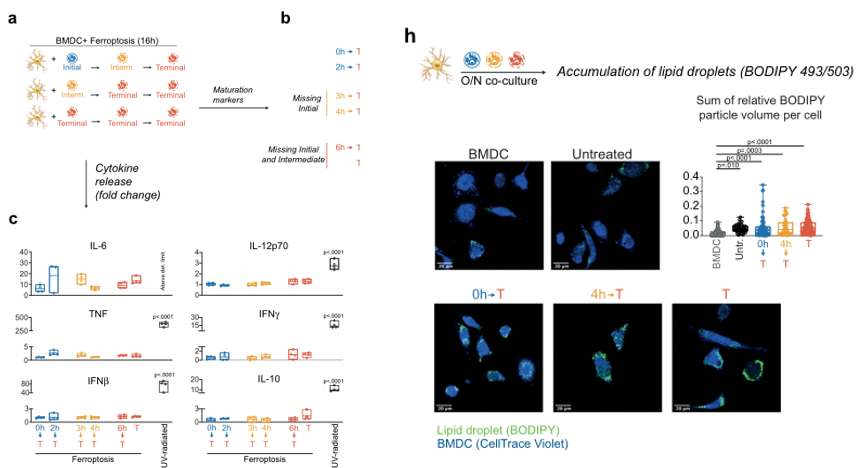
图3(a)将GPX4水平下降、必将出现铁死亡的MCA205细胞与树突状细胞共孵育16 h。为了研究不同铁死亡阶段对树突状细胞成熟的作用,使用了不同细胞死亡阶段的iGPX4KD细胞。所有条件下iGPX4KD细胞在共培养过程中均达到终末期。(c)树突状细胞与铁死亡性细胞孵育后细胞因子产生的分析。(h)用BODIPY 493/503 nm探针对脂滴积累进行显微镜分析。
4. DC吞噬铁死亡的肿瘤细胞会抑制与适应性免疫反应相关基因的表达
为了进一步探讨铁死亡性癌细胞对树突状细胞的影响,作者对吞噬铁死亡性的BMDC进行了转录组学分析。首先将带有荧光的Jurkat细胞进行铁死亡诱导,后跟BMDC细胞进行共培养,通过流式分选的方式,筛选吞噬铁死亡细胞后的DC细胞,进行转录组测序(图4a),通过无偏的全局路径分析,吞噬铁死亡细胞后,DC细胞中与适应性免疫反应相关的基因受到了明显的抑制,如NFKB家族基因,趋化因子和细胞因子等(图4c)。
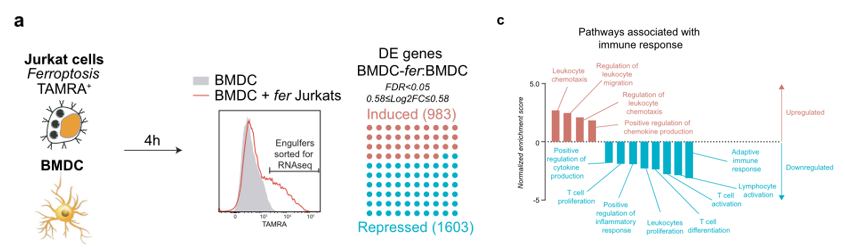
图4 (a)荧光标记的铁死亡性Jurkat细胞与BMDC共培养4小时后,对携带铁死亡性吞噬物的BMDC进行分类并进行小鼠总RNA测序(n = 4个独立收集的样本)。铁死亡性Jurkat细胞导致2586个基因的转录变化。(c)携带铁死亡性货物的BMDC的GSEA通路分析揭示了诱导适应性免疫反应相关通路的转录变化。
5. 铁死亡性细胞损害树突状细胞进行抗原交叉呈递的能力
为了验证BMDC-fer细胞RNAseq的结果,作者从功能上研究了BMDC在暴露于铁死亡性癌细胞后进行抗原交叉呈递的潜力,并诱导抗原特异性ctl的克隆扩增。抗原交叉递呈,指的是MHC 分子对抗原的提呈存在交叉提呈现象,即MHC I 类分子也能提呈外源性抗原,而内源性抗原也能通过MHC II 类途径加以提呈。作者通过DC细胞吞噬铁死亡细胞,并将DC细胞与OT1小鼠(该CD8+T细胞能够特异性识别OVA抗原)的CD8+T细胞进行共孵育(图5a),发现会抑制T细胞的增殖(图5b,c),即铁死亡细胞的外源性抗原抑制DC细胞MHC I对于CD8+T细胞的激活。
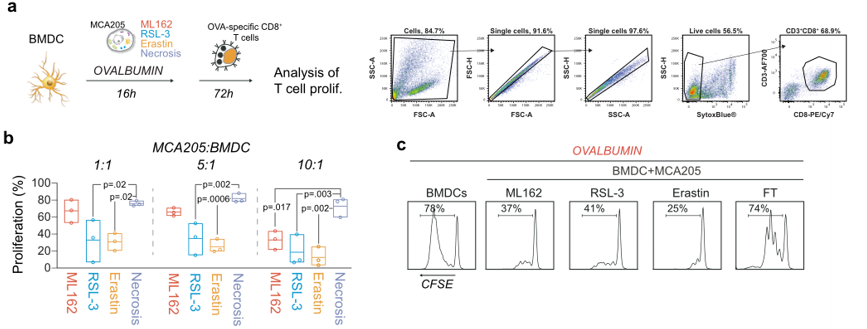
图5 (a)将骨髓来源的树突状细胞(BMDC)与可溶性OVA孵育,培养条件为强铁死亡刺激(ML162 0.5µM,14h;RSL-3,0.5μM, 14 h;Erastin 2.0µM,24h)或坏死癌细胞(3循环冻融)。然后,将荧光标记的ova特异性CD8+细胞加入共培养中,72 h后评估其增殖情况。流式细胞仪点图显示门控策略。(b)与BMDC共孵育铁死亡和坏死细胞后,增殖的ova特异性CD8+ T细胞的百分比。(c)与BMDC暴露于MCA205细胞、铁死亡增多或意外坏死死亡后ova特异性t细胞增殖的代表性直方图。
6. 与凋亡和坏死性凋亡相比,铁死亡对肿瘤生长的控制作用较弱,并降低能够凋亡的免疫原性
接下来,作者使用表达ova的非致瘤性BM1小鼠胚胎成纤维细胞通过凋亡、坏死或铁死亡,在预防接种模型中比较不同细胞死亡方式的免疫原性。首先,作者使用过表达OVA抗原的小鼠胚胎成纤维细胞作为疫苗,先进行免疫反应后再进行皮下成瘤,发现铁死亡与凋亡和坏死相比,几乎完全没有起到预防肿瘤增殖及延长荷瘤小鼠生存期的作用(图6a,b)。于是作者接下来换了一种方式,使用了负载OVA抗原的cDC1细胞进行皮下注射,证明仍然无法对肿瘤生长起到有效影响(图6c)。最后,作者使用米托蒽醌诱导细胞凋亡,发现当加入铁死亡细胞与凋亡细胞进行共同免疫时,能够显著抑制由凋亡细胞疫苗带来的抗肿瘤效果(图6e)。
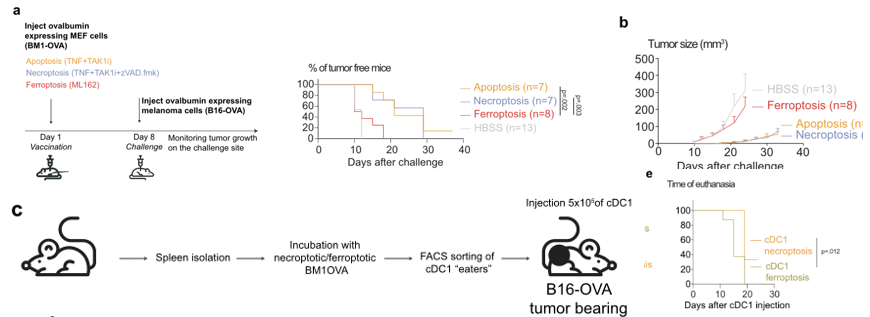
图6 (a)一种预防接种模型,评估铁死亡的免疫原性。(b)接种BM1-OVA细胞后B16-OVA源性黑色素瘤的肿瘤大小。数据以平均值±SEM表示。(c)治疗性疫苗接种实验方案。传统的树突状细胞1型(cDC1)携带铁死亡或坏死物质被皮下注射到黑素瘤小鼠。(e)根据肿瘤大小决定安乐死时间的比较。
研究总结 ✦
这项研究表明癌细胞的铁死亡不是一种免疫原性细胞死亡。铁死亡性癌细胞的这些免疫抑制特性甚至可以推翻凋亡的免疫原性细胞死亡。这项研究的发现对实验和临床免疫治疗具有深远的意义,为未来铁死亡与肿瘤的研究提供了一个明确的认知——单纯的铁死亡会通过抑制免疫微环境中的免疫细胞进行免疫逃逸。
2022年度国自然医学部国自然40大科研热点的中标数统计如下:
2022热点 | 2022中标数 | 2022热点 | 2022中标数 |
免疫调控 | 907 | 中性粒细胞 | 112 |
巨噬细胞 | 591 | 反馈回路 | 104 |
线粒体 | 491 | 乳酸化 | 104 |
血管功能 | 487 | 可变剪接 | 71 |
外泌体 | 470 | AI机器学习 | 67 |
自噬 | 404 | 类器官 | 67 |
铁死亡 | 337 | 炎症小体 | 62 |
干细胞 | 329 | 染色质重塑 | 58 |
代谢重编程 | 325 | 单细胞测序 | 54 |
m6A/m5C/m7G | 320 | 糖基化 | 50 |
泛素化 | 225 | 低氧缺氧 | 50 |
circRNA | 221 | 相分离 | 50 |
lncRNA | 204 | 泛凋亡PANoptosis | 42 |
细胞焦亡 | 175 | 细胞衰老 | 37 |
组蛋白 | 171 | 胞葬 | 33 |
肠道菌群 | 133 | CRISPR | 33 |
乙酰化 | 125 | 增强子 | 29 |
内质网 | 125 | 精氨酸甲基化 | 25 |
转录调控 | 112 | 迁移体 | 8 |
糖酵解 | 112 | 血管拟态 | 8 |
版权声明:本网站所有注明来源“医微客”的文字、图片和音视频资料,版权均属于医微客所有,非经授权,任何媒体、网站或个人不得转载,授权转载时须注明来源:”医微客”。本网所有转载文章系出于传递更多信息之目的,且明确注明来源和作者,转载仅作观点分享,版权归原作者所有。不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。 本站拥有对此声明的最终解释权。































 关注公众号
关注公众号 安卓客户端
安卓客户端










发表评论
注册或登后即可发表评论
登录注册
全部评论(0)