
2022-11-23 来源 : 学术查
肿瘤干细胞(CSCs)是一种静止细胞,具有自我更新能力和多能性,被认为是肿瘤发生、转移、复发的“种子细胞”。CSCs能通过其自我更新和分化为多种细胞亚型的能力产生多种肿瘤,并获得免受大多数传统化疗药物侵害的耐药性。结果便导致这些小亚群持续性细胞形成更具侵袭性和化学耐受性的肿瘤,造成癌症治疗失败和复发。因此,识别和靶向这些相对罕见的癌细胞被认为是对癌症的治疗至关重要的。
在今天要介绍的这项研究中,作者通过单细胞测序鉴定了一种胃癌肿瘤干细胞的新型表面标记物,为胃癌的靶向治疗提供了可能的靶点。
胃癌(GC)是全球死亡数第三的癌症,尽管相关的诊断和治疗策略取得了很多进展,但GC患者的临床和预后效果仍不容乐观。2022年11月14日,济宁医科大学张斌等在Journal of experimental & clinical cancer research : CR(IF=12.658)杂志上发表了题为“AQP5 complements LGR5 to determine the fates of gastric cancer stem cells through regulating ULK1 ubiquitination”的文章。在该研究中,作者检测并对比了胃黏膜组织和胃肿瘤组织的单细胞转录组,并鉴定及表征了一种新型胃癌上皮细胞表面标记物AQP5。作者的研究确定了AQP5通过与E3连接酶TRIM21相互作用并促进关键自噬蛋白ULK1的泛素化来增强胃癌干细胞(GC-CSCs)的自噬,为胃癌的临床治疗提供了新的靶向治疗方案。

原文链接:https://link.springer.com/article/10.1186/s13046-022-02532-w
 研究背景
研究背景
靶向CSCs的治疗策略是癌症治疗的潜在方法,而CSCs主要依赖于细胞表面标志物来区分和鉴定。例如,富含亮氨酸的含有G蛋白偶联受体5(LGR5),也称为G蛋白偶联受体49(GPR49),是一种特征明确的干细胞表面标志物,在多个组织/器官中表达,包括胃、小肠、结肠和肝脏。虽然LGR5在干细胞中起着重要的调节作用,但它在正常干细胞和CSCs中都高表达,这使得在癌症靶向治疗中难以区分正常干细胞和CSC。此外,LGR5在不同来源的肿瘤的CSC中普遍高表达。因此,鉴定能够特异性识别和表征特定起源肿瘤CSC的新型表面标志物仍然是理解肿瘤生物学和开发基于CSC的治疗策略的关键所在。
研究内容
1.AQP5在GC-CSCs中特异性表达
为了探究不同来源肿瘤的CSCs是否具有特异性的表面标记物,作者对胃癌组织和胃黏膜组织进行了单细胞转录组测序,并鉴定出七种类型的细胞(图1a)。测序结果显示GC-CSCs相比于胃癌上皮细胞及胃黏膜组织干细胞(GM-SCs)都有更高表达水平的基因总共有5种,包括aqp5、reg1a、reg3a、ctgf和lyz(图1b-e)。其中,aqp5基因相比于其它4种基因在GC-CSCs中上调程度最高(图1f-h)。另外,作者用qPCR分析了aqp5在肺、肝、肠等其他组织来源的CSCs中的表达情况,发现aqp5只在GC-CSCs中特异性的高表达(图1i)。以上结果表明AQP5可能是GC-CSCs的一种特异性表面标记物。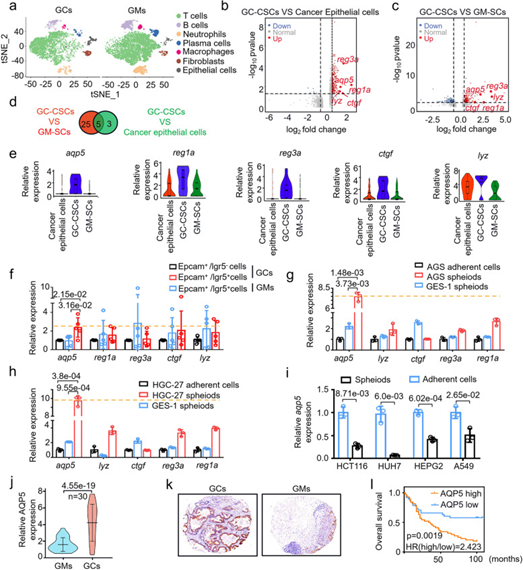
图1.aqp5在GC-CSCs中特异性高表达
2.AQP5在胃癌发生过程中起促进作用
为了研究AQP5对胃癌发生的影响,作者检测了AQP5的mRNA和蛋白水平的表达量,结果表明相比于GMs, AQP5在GCs中表达量显著上调(图1j)。接着,作者通过胃癌组织微阵列评估了AQP5的临床和病理作用,结果显示AQP5的高表达与胃癌患者的存活率低显著相关(图1k-l)。同时,通过对多个胃癌患者进行多元回归分析,作者发现AQP5的表达量与肿瘤的分级呈现正相关(表1)。以上结果表明AQP5在胃癌组织中高表达,并且与临床上胃癌的发生紧密相关。
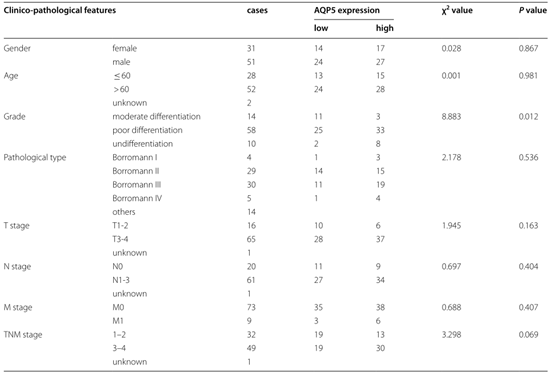
表1.AQP5表达与临床病理特征的相关性
3.AQP5与GC-CSCs经典标记物的生物学关系
此外,作者检测了从胃癌组织中分选和纯化的GC-CSCs中AQP5和LGR5的表达。结果表明AQP5阳性胃癌细胞的LGR5的表达量均较高,而AQP5和CD133在LGR5阳性的GC细胞和胃粘膜细胞中表达水平相同(图2f-j)。以上结果表明AQP5与经典的肿瘤干细胞标记物密切相关,其和LGR5/CD133在胃癌组织中共同表达。
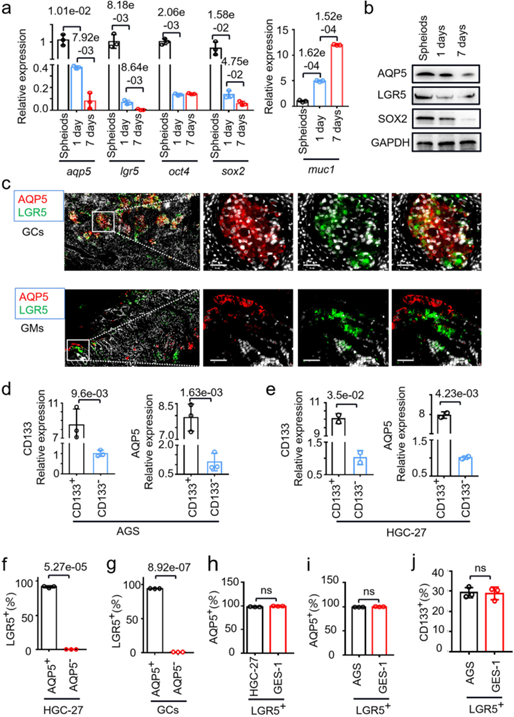
图2.AQP5表达与干性标记物表达的相关性
4.AQP5促进GC-CSCs的自我更新和肿瘤发生
接下里,作者检测了AQP5对GC-CSCs生物学功能的影响。与预期结果一致,AQP5敲除后GC-CSCs的成球显著降低,反之AQP5过表达后GC-CSCs的成球显著增强(图3a-b)。为了进一步检测AQP5对GC-CSCs干性的影响,作者检测了干细胞标记物的表达情况。结果发现AQP5敲除后干细胞标记物LGR5和SOX2的表达量显著降低,且CD44/CD133/LGR5阳性细胞的比例也有所下降,反之过表达后趋势相反(图3c-g)。随后,作者通过皮下注射不同浓度的GCs到免疫缺陷小鼠中构建异种移植模型。结果发现AQP5敲除后肿瘤的形成率及肿瘤的重量和体积都显著下降,而过表达后趋势相反(图3h-m)。以上结果表明AQP5在体外水平促进GCs的干性维持并在体内水平促进肿瘤的发生和生长。
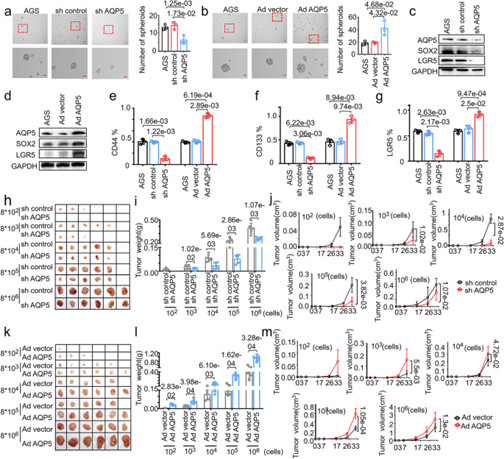
图3. AQP5在体内外促进GC-CSCs的肿瘤生物学功能
5.AQP5和LGR5在GC-CSCs致瘤中起互补作用
基于AQP5和LGR5在GCs中共表达,作者将两者分别单独敲除或者共同敲除来研究两者的关系。有趣的是,将AQP5和LGR5同时敲除后能够显著抑制GCs的成球和迁移,然而两者单独敲除所造成的效果则不明显(图4a-b)。同样,在异种移植模型中,两者共同敲除的GCs的肿瘤形成相比单独敲除的GCs受到了更加明显的抑制作用(图4c-e)。以上结果表明AQP5和LGR5其协同作用以促进GC-CSCs的肿瘤形成。
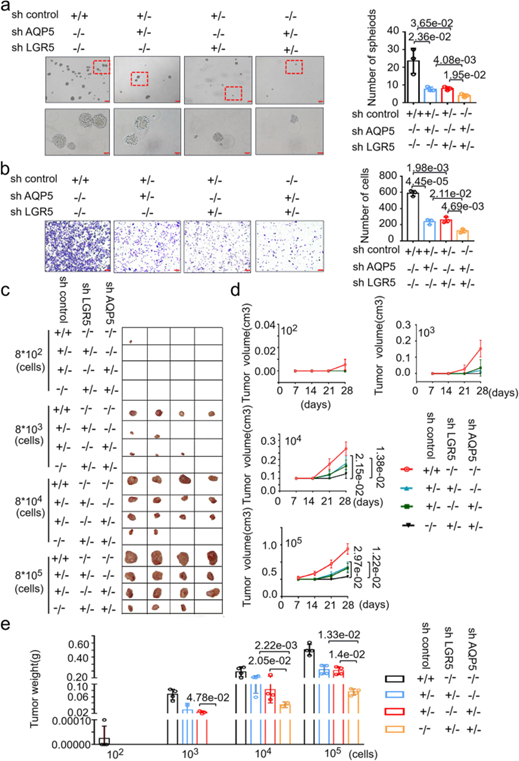
图4. AQP5和LGR5共同敲除显著抑制GC-CSCs的肿瘤生物学功能
6.AQP5激活GC-CSCs的自噬
AQP3是与AQP5同蛋白家族的成员,其已被报道能通过激活自噬促进肿瘤发生,因此作者推测AQP5可能也是通过调控姿势来决定GC-CSCs的命运。AQP5敲除后,LC3II的表达下调而P62表达上调;AQP5 过表达后趋势则相反,并且会促进LC3自噬小体的形成(图5a-e)。接下来,作者将AQP5过表达GCs的自噬关键蛋白ATG7敲除,结果发现ATG7敲除显著抑制了由AQP5引起的成球和干性标记物SOX2的表达(图5f-i)。同时,使用自噬抑制剂氯喹啉CQ也能抑制LC3II的表达和GCs的成球(图5j-l)。以上结果表明AQP5通过激活自噬来调控GC-CSCs的功能和干性。

图5. AQP5通过激活自噬促进GC-CSCs的功能
7.AQP5通过ULK1泛素化促进GC-CSCs的功能
鉴于泛素化涉及自噬调控的多个方面,作者推测AQP5通过调控关键自噬蛋白的泛素化来促进GC-CSCs的自噬激活。AQP5过表达后,ULK1而不是其他的关键自噬蛋白发生了显著的泛素化,而且是通过63号赖氨酸(K63)被泛素分子修饰来完成的,而AQP5敲除后则趋势相反(图6a-d)。同时,将GC-CSCs的内源性泛素敲除后,由K63介导的ULK1泛素化受到了显著抑制;且K63突变后的泛素化也无法引起由AQP5过表达引起的干性标记物表达上调(图6e-f)。
TRIM21是一种AQP5结合蛋白,并且是一种与泛素化紧密相关的E3泛素连接酶。TRIM21敲除后,AQP5引起的K63介导的ULK1泛素化、GC-CSCs的成球、干性标记物及自噬关键蛋白的表达都受到了抑制(图6g-h)。而AQP5对TRIM21的表达量并无影响,而是促进ULK1和TRIM21的互作(图6h-k)。以上结果表明AQP5通过募集E3泛素连接酶到ULK1中促进K63介导的ULK1泛素化。

图6. AQP5通过TRIM21促进ULK1泛素化
研究总结
在本研究中,作者对比了胃粘膜组织和胃肿瘤组织的单细胞转录组,并鉴定了一种GC-CSCs的新型特异性表面标记物AQP5。结果表明,AQP5与LGR5协同作用,通过募集E3泛素连接酶TRIM21与关键自噬蛋白ULK1互作,引起K63介导的ULK1泛素化来增强GC-CSCs的自噬,最终导致GC-CSCs的干性增强并促进肿瘤的发生。
此研究确定了胃癌肿瘤干细胞的特异性表面标记物,为人类胃癌的临床治疗提供了新的研究思路,有望转化为有效的靶向治疗。
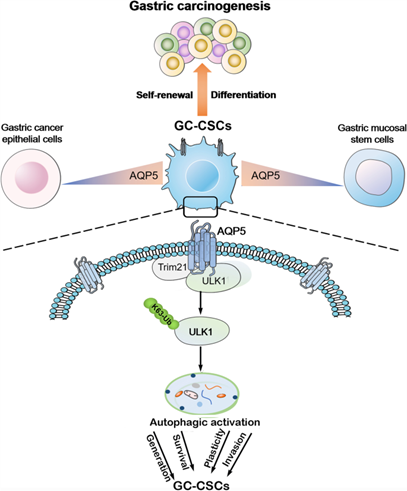
机制模式图
2022年度国自然医学部国自然40大科研热点的中标数统计如下:
2022热点 | 2022中标数 | 2022热点 | 2022中标数 |
免疫调控 | 907 | 中性粒细胞 | 112 |
巨噬细胞 | 591 | 反馈回路 | 104 |
线粒体 | 491 | 乳酸化 | 104 |
血管功能 | 487 | 可变剪接 | 71 |
外泌体 | 470 | AI机器学习 | 67 |
自噬 | 404 | 类器官 | 67 |
铁死亡 | 337 | 炎症小体 | 62 |
干细胞 | 329 | 染色质重塑 | 58 |
代谢重编程 | 325 | 单细胞测序 | 54 |
m6A/m5C/m7G | 320 | 糖基化 | 50 |
泛素化 | 225 | 低氧缺氧 | 50 |
circRNA | 221 | 相分离 | 50 |
lncRNA | 204 | 泛凋亡PANoptosis | 42 |
细胞焦亡 | 175 | 细胞衰老 | 37 |
组蛋白 | 171 | 胞葬 | 33 |
肠道菌群 | 133 | CRISPR | 33 |
乙酰化 | 125 | 增强子 | 29 |
内质网 | 125 | 精氨酸甲基化 | 25 |
转录调控 | 112 | 迁移体 | 8 |
糖酵解 | 112 | 血管拟态 | 8 |
版权声明:本网站所有注明来源“医微客”的文字、图片和音视频资料,版权均属于医微客所有,非经授权,任何媒体、网站或个人不得转载,授权转载时须注明来源:”医微客”。本网所有转载文章系出于传递更多信息之目的,且明确注明来源和作者,转载仅作观点分享,版权归原作者所有。不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。 本站拥有对此声明的最终解释权。


































 关注公众号
关注公众号 安卓客户端
安卓客户端










发表评论
注册或登后即可发表评论
登录注册
全部评论(0)